ė╬ļx░▒š{┐žī”╬█─ÓĖ▀║¼╣╠ģÆ觎¹╗»Ę┤æ¬Ų„Ą─ė░Ēæ
ųąć°╬█╦«╠Ä└Ē╣ż│╠ŠW ĢrķgŻ║2017-2-11 8:03:27
╬█╦«╠Ä└Ē╝╝ąg | ģRŠ█╚½Ū“Łh▒Ż┴”┴┐Ż¼ĮĄĄ═Ų¾śIų╬╬█│╔▒Š
ĪĪĪĪĖ▀║¼╣╠[▀M┴Ž║¼╣╠┬╩(total solidsŻ¼TS)>10%]ģÆ觎¹╗»╩ŪĮŌøQ╬ęć°├ō╦«╬█─Ó╠Ä└Ē╠Äų├å¢Ņ}Ą─ųžę¬═ŠÅĮ.ŽÓ▒╚é„ĮyĄ─Ą═║¼╣╠ģÆ觎¹╗»(║¼╣╠┬╩×ķ1%~5%)[1]Ż¼Ė▀║¼╣╠ģÆ觎¹╗»╝╝ąg╔§ų┴┐╔ęįų▒Įė└¹ė├╬█╦«╠Ä└ĒÅSĄ─├ō╦«╬█─Ó(║¼╣╠┬╩×ķ20%)Ż¼ę“Č°Š▀ėąįO╩®¾wĘeąĪĪó╦«║─╝░─▄║─▌^Ą═Ą╚ā×ä▌.┴Ē═ŌŻ¼Ė▀║¼╣╠ģÆ觎¹╗»ŽĄĮy║═é„ĮyĄ─Ą═║¼╣╠ģÆ觎¹╗»ŽĄĮyŽÓ▒╚Ż¼VSĮĄĮŌ┬╩╗∙▒ŠŽÓ«öŻ¼Č°Ė▀║¼╣╠ģÆ觎¹╗»ŽĄĮyĄ─å╬╬╗╚▌Ęe«aÜŌ┬╩Ė³Ė▀[2]Ż¼Š▀ėąĖ³║├Ą─æ¬ė├Ū░Š░.
ĪĪĪĪ╚╗Č°Ż¼ė╔ė┌╬█─ÓĖ▀║¼╣╠ģÆ觎¹╗»ŽĄĮyĄ─▀M┴Žųą║¼Ą¬╬’┘|╚ńĄ░░ū┘|Īó─“╦žĄ╚║¼┴┐Ė³Ė▀Ż¼Įø▀^╬ó╔·╬’Ą─ū„ė├Ż¼╦³éāĢ■ūŅĮKĮĄĮŌ×ķąĪĘųūėĄ─░▒Ą¬▓ó└█Ęeį┌ŽĄĮyųą.▀^Ė▀Ą─░▒Ą¬ØŌČ╚Ģ■ī”╬ó╔·╬’«a╔·ČŠ║”ū„ė├[3]Ż¼Å─Č°ĮĄĄ═š¹éĆŽ¹╗»ŽĄĮyĄ─«aÜŌąį─▄.┐é░▒Ą¬(TAN)ėųĘų×ķõ@Ė∙ļxūė║═ė╬ļxæBĄ─░▒(FAN)Ż¼Ųõųąė╬ļx░▒Š▀ėą▌^Ė▀Ą─╝Ü░¹─żØB═ĖąįČ°▒╗šJ×ķ╩Ū«a╔·░▒ęųųŲĄ─ų„ę¬įŁę“[4].╬ó╔·╬’▒╗░▒Ą¬ęųųŲĄ─┐╔─▄ÖCųŲ╩Ūė╬ļx░▒ū„×ķ╩Ķ╦«ąįĘųūėŻ¼═©▀^▒╗äėöU╔óū„ė├▀M╚ļ╝Ü░¹Ż¼ŲŲē─┴╦╝Ü░¹ā╚═Ō┘|ūėŲĮ║Ō▓óī¦ų┬ŌøĄ─╚▒Ę”[5].┴Ē═ŌŻ¼▀M╚ļ╝Ü░¹Ą─ė╬ļx░▒į┌╝Ü░¹ā╚▐Dūā×ķõ@Ż¼õ@į┌╝Ü░¹ā╚Ęe└█Ė─ūā┴╦╝Ü░¹ā╚Ą─pHŻ¼Å─Č°ī”╝Ü░¹«a╔·ČŠ║”ū„ė├[6].ĻPė┌ė╬ļx░▒ī”ģÆ觎¹╗»ŽĄĮyĄ─ė░ĒæŻ¼ė╔ė┌ŽĄĮyĄ─║¼╣╠┬╩Īó╗∙┘|Īó£žČ╚║═ĮėĘN─Ó±Z╗»│╠Č╚Ą╚Ė„ĘNę“╦žĄ─▓╗═¼Ż¼╦∙Ą├│÷Ą─ė╬ļx░▒ęųųŲķōųĄę▓Å─Äū╩«ĄĮÄū░┘║┴┐╦╔²▓╗Ą╚[7].└²╚ńŻ¼é„ĮyĄ─Ą═║¼╣╠ģÆ觎¹╗»ŽĄĮyųąŻ¼FANį┌145 mgĪżL-1Ż¼pH×ķ7.43Ģr▒ŃĢ■╩╣╝ęŪ▌╝S▒ŃæęĖĪę║ģÆ觎¹╗»Ą─«aÜŌ┴┐Ž┬ĮĄ27%[8].į┌Ė▀║¼╣╠ģÆ觎¹╗»ŽĄĮyųąŻ¼HidakaĄ╚[9]Ą─īŹ“×░l¼FFANį┌▓╗ĄĮ600 mgĪżL-1Ą─Śl╝■Ž┬Ż¼Ė▀║¼╣╠ŽĄĮyCODĮĄĮŌ┬╩Š═╩▄ĄĮ├„’@ęųųŲ.╚╗Č°Ż¼╬ó╔·╬’ī”ė┌ė╬ļx░▒Ą──═╩▄Č╚┐╔ęįĖ³Ė▀Ż¼KimĄ╚[10]░l¼FFAN×ķ700 mgĪżL-1ĢrŽĄĮy╚į─▄ĘĆČ©▀\ąą▓╗╩▄ęųųŲ.Č°CalliĄ╚[11]└¹ė├╚╦╣ż┼õ╦«ū„×ķ╗∙┘|Ż¼į┌╠ßĖ▀ė╬ļx░▒Ą─▀^│╠ųąŻ¼CODĮĄĮŌ┬╩ę╗ų▒ŠS│ųį┌78%~96%Ż¼╝┤╩╣į┌FANūŅĮK▀_ĄĮ800 mgĪżL-1Ģrę└╚╗ŠS│ųį┌80%Ż¼ŽĄĮyąį─▄▓óø]ėą╩▄ĄĮęųųŲ.╦∙ęįŻ¼ģÆ觎¹╗»░▒ęųųŲ蹊┐Ą─ųž³c▓╗āHį┌ė┌½@Ą├ė╬ļx░▒Ą─ęųųŲķōųĄØŌČ╚Ż¼▀Ćį┌ė┌蹊┐ė╬ļx░▒ØŌČ╚Ė─ūā▒│║¾ŽÓĻPĄ─╬ó╔·╬’ĘN╚║ĮYśŗ║═┤·ųx═ŠÅĮĄ─ūā╗»Ż¼Å─Č°šµš²Įę╩Š░▒ęųųŲĄ─ÖCųŲ.ė╬ļx░▒Ą─ėŗ╦ŃĘĮĘ©┼c┐é░▒Ą¬ĪópH║═£žČ╚ėąĻPŻ¼į┌£žČ╚▓╗ūāĄ─ŪķørŽ┬Ż¼ŁhŠ│ųąĄ─pHųĄįĮĄ═Ż¼FANę▓įĮĄ═.└²╚ńį┌ģÆ觎¹╗»£žČ╚×ķ37ĪµĄ─Śl╝■Ž┬Ż¼«öpHÅ─8.0ĮĄĄ═ų┴7.0ĢrŻ¼FANš╝TANĄ─▒╚└²īóĢ■Å─11%Ž┬ĮĄų┴1.2%Ż¼ę“Č°ĮĄĄ═pH╩Ūę╗ĘNĮĄĄ═ŽĄĮyųąFANØŌČ╚Ą─ĘĮĘ©.▓╗═¼Ą─╬ó╔·╬’Ą─▀mę╦pHĘČć·ę▓ėą╦∙▓╗═¼, «a╝ū═ķŠ·Ą─ūŅ▀mę╦pHĘČć·×ķ6.5~7.8[12]Ż¼Č°«a╦ߊ·ūŅ╝čpHį┌5.5~7.0[3]Ż¼┐žųŲpHį┌╬ó╔·╬’▀mę╦Ą─ĘČć·ā╚┐╔─▄┐╔ęį£p▌pFANĄ─ęųųŲū„ė├[9].
ĪĪĪĪ▒ŠīŹ“×▓╔ė├9 LĄ─░ļ▀B└mģÆ觎¹╗»čbų├Ż¼ęį├ō╦«╬█─Ó×ķįŁ┴ŽŻ¼Ęųäe═©▀^ĮĄĄ═pH║═╚╦×ķ═Č╝ė░▒Ą¬Ą─╩ųČ╬üĒĮĄĄ═║═╠ßĖ▀Ę┤æ¬Ų„ā╚Ą─ė╬ļx░▒ØŌČ╚Ż¼ęį┐╝▓ņā╔ĘNš{┐ž╩ųČ╬Ž┬ė╬ļx░▒ØŌČ╚Ą─ūā╗»ī”ģÆ觎¹╗»ŽĄĮyąį─▄Ą─ė░Ēæ.┼c┤╦═¼ĢrŻ¼└¹ė├Ė▀═©┴┐£yą“║═Č©┴┐PCRĄ─Ęų╬÷ĘĮĘ©Ż¼čąŠ┐Ė▀║¼╣╠╬█─ÓģÆ觎¹╗»ŽĄĮyį┌▓╗═¼Ą─ė╬ļx░▒š{┐ž▓▀┬įŽ┬ąį─▄ģóöĄęį╝░ŽÓæ¬Ą─╝ÜŠ·║═╣┼Š·ĘN╚║ĮYśŗ║═öĄ┴┐Ą─ūā╗»Ż¼ęįŲ┌×ķūŅĮK║Ž└Ē┐žųŲĖ▀║¼╣╠ģÆ觎¹╗»¾wŽĄųąė╬ļx░▒ØŌČ╚Ż¼▒ŻūCŽ¹╗»ŽĄĮy┴╝║├ĘĆČ©Ąž▀\ąą╠ß╣®└Ēšōų¦ō╬.
ĪĪĪĪ1 ▓─┴Ž┼cĘĮĘ©
ĪĪĪĪ1.1 ģÆ觎¹╗»īŹ“×
ĪĪĪĪīŹ“×ųąĄ─├ō╦«╬█─Ó╚Īūį╔Ž║Ż─│╬█╦«╠Ä└ĒÅSĄ─├ō╦«╬█─ÓÖCĘ┐Ż¼├ō╦«║¾Ą─╬█─Ó║¼╣╠┬╩(TS)║═ō]░ląį╣╠¾w(VS)š╝┐é╣╠¾w▒╚└²(VS/TS)Ęųäe×ķ20.1%║═53.9%.īŹ“×╦∙ė├ĮėĘN╬█─Ó╚ĪūįīŹ“×╩ęųąĘĆČ©▀\ąąĄ─├ō╦«╬█─Óųą£žģÆ觎¹╗»Ę┤æ¬Ų„Ż¼TS║═VS/TSĘųäe×ķ13.2%║═46.0%.▀M┴Ž├ō╦«╬█─Ó║═ĮėĘN─ÓĄ─╠╝Ą¬▒╚(C/N)Ęųäe×ķ6.9Ż║1║═5.2Ż║1.
ĪĪĪĪ░ļ▀B└m╩ĮĘ┤æ¬Ų„Ą─ėąą¦╚▌Ęe×ķ9 LŻ¼═©▀^┬▌Ħ╩Įöć░Ķčbų├▀Mąąöć░ĶŻ¼ā╚ų├Ą─╦«įĪ£ž┐žčbų├─▄ŠS│ųĘ┤æ¬Ų„ā╚╬’┴Ž£žČ╚×ķ(37Ī└1)Īµ.├┐╚š«aÜŌ┴┐═©▀^ر╩ĮÜŌ¾w┴„┴┐ėŗ£yČ©.Ę┤æ¬Ų„Ą─▐D╦┘×ķ60 rĪżmin-1Ż¼ęį▐Däė/═Żų╣×ķ10 min/10 minĘĮ╩ĮĮ╗╠µ▀\ąą.▀\ąąĄ─╩ū╚šŽ“Ę┤æ¬Ų„ā╚╝ė╚ļ4 kgĮėĘN─Ó║═1 kg╗∙┘|įŁ─ÓŻ¼ų«║¾▀B└m4 d├┐╠ņ╠Ē╝ė1 kg╩┬Ž╚═©▀^ūįüĒ╦«┼õųŲ║├Ą─║¼╣╠┬╩×ķ15%Ą─├ō╦«╬█─Óų┴Ę┤款wŽĄĄ─┐é¾wĘe▀_ĄĮ9 LŻ¼Å─Ą┌6 dķ_╩╝░┤šš╣╠¾w═Ż┴¶Ģrķg(solid retention timeŻ¼SRT)×ķ20 dęį░ļ▀B└mĘĮ╩Į▀M│÷┴Ž(├┐╠ņ▀M│÷┴ŽĖ„ę╗┤╬).
ĪĪĪĪ▒ŠīŹ“×╣▓▀\ąą2┼_Ę┤æ¬Ų„(R1║═R2). R1į┌ĘĆČ©▀\ąąų┴35 d║¾├┐╠ņīó▀m┴┐Ą─ØŌHCl┼c▀M┴Ž╗ņ║ŽŻ¼į┌Ą┌50 dĘ┤æ¬Ų„pHŽ┬ĮĄų┴7.09Ī└0.02Ż¼ļS║¾ĘĆČ©įōpHųĄ41 dŻ¼Ą┌92 d═Żų╣╠Ē╝ėHClų▒ų┴īŹ“×ĮY╩°. R2į┌ĘĆČ©▀\ąąų┴56 d║¾Ż¼ķ_╩╝į┌▀M┴Žųą╠Ē╝ė30 g NH4ClŻ¼▓óę└ō■TANĘų╬÷ĮY╣¹Ęųäeė┌Ą┌58 d║═Ą┌67 d╝ė╚ļ13 g║═35 g NH4Clęį╩╣Ą├Ę┤æ¬Ų„ā╚Ą─TANÅ─3 284 mgĪżL-1į÷╝ėĄĮ5 500 mgĪżL-1.Å─Ą┌70 dķ_╩╝Ż¼×ķ┴╦ŠS│ųTANį┌5 000~6 000 mgĪżL-1Ż¼Ė∙ō■│÷┴Žųą░▒Ą¬Ą─┴„╩¦┴┐ėŗ╦ŃŻ¼├┐╠ņį┌▀M┴Žųą╝ė╚ļ5 g NH4Clų┴īŹ“×ĮY╩°. 70~124 d▀@ę╗ļAČ╬FANÅ─ĘĆČ©ļAČ╬Ą─(400Ī└173) mgĪżL-1╠ßĖ▀ĄĮ┴╦(526Ī└25) mgĪżL-1.ŽĄĮyFANØŌČ╚ĘČć·ūŅĮKĘĆČ©į┌450~600 mgĪżL-1(┐é░▒Ą¬ØŌČ╚×ķ5 000~6 000 mgĪżL-1)ų┴īŹ“×ĮY╩°.
ĪĪĪĪ1.2 ųĖś╦Ęų╬÷ĘĮĘ©
ĪĪĪĪŲĮŠ∙├┐ų▄▓╔╝»ā╔┤╬Ę┤æ¬Ų„Ą─│÷┴Ž▀MąąĘų╬÷Ż¼£yČ©pHĪóTSĪóVS/TSŻ¼ō]░ląįų¼ĘŠ╦ß(volatile fatty acidsŻ¼VFAs)ØŌČ╚Īó┐éēAČ╚Īó┐é░▒Ą¬(TAN)ØŌČ╚ĪóÜŌ¾w│╔Ęų.├┐╠ņėøõø«aÜŌ┴┐. pH£yČ©▓╔ė├īŹ“×╩ęė├pHėŗ(S210, METTLER, Switzerland).Ž¹╗»╬█─ÓśėŲĘį┌13 000 rĪżmin-1Ž┬ļxą─20 min║¾▀Mąą▀^×VŻ¼×Vę║Įø╬ó┐ū×V─ż(0.45 ”╠m)▀^×V║¾Ż¼ŽĪßīę╗Č©▒ČöĄ▓ó╝ė╚ļ╝ū╦ßš{š¹pHĪ▄2.0Ż¼į┘╩╣ė├ÜŌŽÓ╔½ūV[2010 plus, Shimadzu, JapanŻ¼╗čµļxūė╗»Öz£yŲ„(FID)]£yČ©VFAØŌČ╚.╦∙╩╣ė├Ą─╔½ūVų∙×ķRtx-WAXą═├½╝Ü╣▄ų∙(30 mĪ┴0.25 mmĪ┴0.25 ”╠m). TSĪóVS/TSĪó┐éēAČ╚║═TANØŌČ╚▓╔ė├ś╦£╩ĘĮĘ©£yĄ├[13].šėÜŌųą╝ū═ķ║═Č■č§╗»╠╝Ą─║¼┴┐£yČ©▓╔ė├čbėą¤ßī¦Öz£yŲ„(TCD)Ą─ÜŌŽÓ╔½ūVāx(GC112A, INESA, China)Ż¼╔½ūVų∙×ķGDX-102(2 mĪ┴4 mm).ė╬ļx░▒ØŌČ╚Ė∙ō■┐é░▒Ą¬ØŌČ╚ĪópH║═£žČ╚ėŗ╦ŃĄ├ĄĮ[14], Š▀¾wėŗ╦Ń╣½╩Į╚ńŽ┬Ż║

ĪĪĪĪ(1)
ĪĪĪĪ╩ĮųąŻ¼[FAN]║═[TAN]Ęųäe×ķė╬ļx░▒║═┐é░▒Ą¬Ą─ØŌČ╚(mgĪżL-1);T×ķ¤ß┴”īW£žČ╚(K).
ĪĪĪĪVSĮĄĮŌ┬╩(VSreduction)ė╔╩Į(2)[15]ėŗ╦ŃĄ├│÷(╝┘Č©ŽĄĮyųą¤oÖCĮMĘųŻ¼╝┤▓╗─▄▒╗ĮĄĮŌĄ─╬’┘|Ą─┴┐▒Ż│ų║ŃČ©)Ż║
ĪĪĪĪ(2)
ĪĪĪĪ╩Įųą, VSdigestate║═VSfeedĘųäe×ķģÆ觎¹╗»║¾Ą─šėį³║═▀M┴Ž╬█─ÓųąĄ─VSš╝TSĄ─▒╚└²(VS/TSŻ¼%).
ĪĪĪĪ1.3 ╬ó╔·╬’Ęų╬÷
ĪĪĪĪ1.3.1 ╬ó╔·╬’ĘN╚║ĮYśŗĘų╬÷
ĪĪĪĪČ©Ų┌▒Ż┴¶Ę┤æ¬Ų„│÷┴Žė┌2 mLā÷┤µ╣▄ųąŻ¼ė┌-80Īµ└õā÷▒Ż┤µ. DNAśėŲĘ╩╣ė├═┴╚└DNA╠ß╚Īįćä®║ą(PowerSoilŻ¼MO BIO, USA)╠ß╚Ī.Ęų╬÷ųą▀xė├Ą─8éĆR1śėŲĘüĒūįĄ┌28Īó35Īó42Īó49Īó77Īó88Īó109║═124 dŻ¼┤·▒Ē┴╦ĘĆČ©Īóš{┐žĪóęųųŲĘĆČ©║═╗ųÅ═▀@4éĆļAČ╬.╦∙▀xė├Ą─16éĆR2śėŲĘüĒūįĄ┌4Īó10Īó16Īó21Īó28Īó35Īó42Īó49Īó56Īó58Īó63Īó70Īó77Īó84Īó91║═102 dŻ¼┤·▒Ē┴╦R2åóäėĪóĘĆČ©Īó╠ßĖ▀ė╬ļx░▒ĪóĖ▀ė╬ļx░▒ęųųŲĘĆČ©▀@4éĆļAČ╬.╠ß╚Ī│÷Ą─DNAĮø▀^PCRöUį÷Ż¼▓╔ė├Ė▀═©┴┐£yą“(MiSeq4000, Illumina)Ęų╬÷Ųõųą╣┼Š·┼c╝ÜŠ·Ą─ĘN╚║ĮYśŗ. PCR«a╬’Ž╚└¹ė├QuantiFLuorTMŽĄĮy(Promega)£yČ©ØŌČ╚Ż¼ļS║¾ę└ō■AxyPrep DNAįćä®║ą(AXYGEN, USA)Ą──²─z╗ž╩šĄ─ĘĮĘ©▀Mąą╠ß╝ā.╠ß╝āĄ─PCR«a╬’Ą─┘|┴┐═©▀^─²─zļŖėŠ┤_Č©.ę²╬’įOėŗĖ∙ō■Illumina╣½╦Š(San Diego, California, USA)Ą─▓┘ū„╩ųāįŻ¼ßśī”╝ÜŠ·║═╣┼Š·Ą─16S rRNA╗∙ę“Ą─═©ė├öUį÷ę²╬’ī”Ęųäe×ķ338f (5Īõ-ACTCCTACGGGAGGCAGCA-3Īõ)/806r (5Īõ-GGA CTACHVGGGTWTCTAAT-3Īõ)[16]║═344f (5Īõ-ACGGG GYGCAGCAGGCGCGA-3Īõ)/915r (5Īõ-GTGCTCCCCC GCCAATTCCT-3Īõ)[17].
ĪĪĪĪ1.3.2 ╝ū═ķŠ·Č©┴┐PCRĘų╬÷
ĪĪĪĪ▒ŠčąŠ┐ųąĖ∙ō■ČÓśėąįĘų╬÷ĮY╣¹Ż¼▀x╚Ī«a╝ū═ķŠ·ųąų„ꬊ·╚║╝ū═ķ░╦»BŪ“Š·┐Ų(Methanosarcinaceae)Ż¼ī”Ųõ16S rRNA╗∙ę“ØŌČ╚ūā╗»▀Mąą┴╦Č©┴┐Ęų╬÷.Č©┴┐PCRīŹ“×▓╔ė├Sybergreen¤╔╣Ō╚Š┴ŽĘ©Ż¼ę²╬’įOėŗ▓╔ė├╝ū═ķ░╦»BŪ“Š·┐Ų16S rRNA╗∙ę“Ą─╠ž«Éę²╬’ī”Ż║Ū░ę²╬’(5Īõ-GAAACCGYGATAAGGGGA-3Īõ)║═║¾ę²╬’(5Īõ-TAGCGARCATCGTTTACG-3Īõ)[18].╩╣ė├Č©┴┐PCRāx(7500 Real Time PCR SystemŻ¼Applied Biosystems)ī”Methanosarcinaceae▀MąąČ©┴┐Ęų╬÷. SybergreenČ©┴┐PCRĘ┤款wŽĄ×ķ10 ”╠L SYBR Premix Ex Taqó“ (Tli RNaseH Plus) (2x) (TaKaRa, Japan), 0.8 ”╠LŪ░ę²╬’(10 ”╠molĪżL-1), 0.8 ”╠L║¾ę²╬’(10 ”╠molĪżL-1), 0.4 ”╠L ROXó“Ż¼2 ”╠L─Ż░ÕDNA║═PCRĄ╚╝ē¤oŠ·╦«Ż¼ūŅĮK¾wĘe×ķ20 ”╠L.├┐éĆśėŲĘ3éĆŲĮąą.Ę┤æ¬▓Į¾EŻ║Ž╚95ĪµŽ┬ŅAūāąį30 sŻ¼╚╗║¾Įø▀^95ĪµŽ┬5 s, 55ĪµŽ┬30 s║═72ĪµŽ┬40 s裣h40┤╬.├┐ę╗┤╬裣hĄ─ūŅ║¾ę╗éĆ▓Į¾EÖz£y¤╔╣ŌųĄ.
ĪĪĪĪ2 ĮY╣¹┼cėæšō2.1 ė╬ļx░▒š{┐žī”ŽĄĮyąį─▄Ą─ė░Ēæ2.1.1 ĮĄĄ═ė╬ļx░▒ØŌČ╚ī”«aÜŌ┴┐║═VSĮĄĮŌ┬╩Ą─ė░Ēæ
ĪĪĪĪR1Ą─ŽÓĻP└Ē╗»ųĖś╦ūā╗»Ū·ŠĆ╚ńłD 1╦∙╩Š.į┌Ę┤æ¬Ų„åóäėļAČ╬(0~24 d)Ż¼╚š«aÜŌ┴┐▓©äė▌^┤¾Ż¼▓óŪęį┌Ą┌13 d║═Ą┌24 d│÷¼Fā╔éĆ«aÜŌĖ▀ĘÕ.įōļAČ╬ėą┤¾┴┐Ą─VFAĘe└█Ż¼TAN║¼┴┐ę▓║▄Ė▀Ż¼šf├„åóäėļAČ╬╦«ĮŌ║═«a╦ߊ·╗ŅąįŽÓī”ė┌«a╝ū═ķŠ·╗ŅąįĖ³Ė▀Ż¼─▄ē“┐ņ╦┘ĮĄĮŌ┤¾┴┐Ą─┤¾ĘųūėėąÖC╬’▓ó╔·│╔VFAŻ¼Č°┤╦Ģr«a╝ū═ķŠ·ė╔ė┌Ųõ╩└┤·ų▄Ų┌ŽÓī”▌^ķLŻ¼▀Ć▓╗─▄┤¾┴┐į÷ų│▓ó╝░ĢrŽ¹║─ęę╦߯¼ī¦ų┬VFAĘe└█║═«aÜŌäĪ┴ę▓©äė. 25 d║¾Ż¼R1Ą─«aÜŌ┴┐ųØuĘĆČ©Ż¼pHę▓ĘĆČ©į┌7.98Ī└0.04Ż¼ĘĆČ©ļAČ╬(25~35 d) FAN╝s×ķ(329Ī└40) mgĪżL-1. BraunĄ╚[19]└¹ė├ę║æBĄ─ži╝S▀MąąģÆč§░lĮ═Ż¼╠ß│÷FANį┌150 mgĪżL-1ĢrŽĄĮyŠ═ķ_╩╝╩▄ĄĮęųųŲŻ¼╦¹éāĄ─Ę┤欎ĄĮyį┌ĮĄĄ═pHų«║¾½@Ą├┴╦Ė³║├Ą─«aÜŌ┴┐.═¼śėŻ¼ZeemanĄ╚[20]į┌┼Ż╝SĄ─Ė▀£žģÆ觎¹╗»▀^│╠ųąŻ¼TAN×ķ3 000 mgĪżL-1Ż¼░l¼Fį┌pH×ķ7.5ĢrŻ¼ąį─▄║▄▓ŅŻ¼Č°«ö╦¹éāīópHĮĄĄĮ7.0║¾Ż¼ė╔ė┌FANĄ─ĮĄĄ═Ż¼╝ū═ķĄ─«a┴┐į÷ķL┴╦4▒Č.ę“┤╦Ż¼×ķ┴╦╩╣Ą├ė╬ļx░▒ØŌČ╚Ž┬ĮĄŻ¼£p▌p╬ó╔·╬’╦∙╩▄Ą─ė╬ļx░▒Ą─ęųųŲū„ė├Ż¼½@Ą├Ė³║├Ą─«aÜŌąį─▄Ż¼Å─Ą┌35 dķ_╩╝╚╦×ķĄžŽ“R1ųą╠Ē╝ėHClŻ¼╩╣Ą├pHųØuŽ┬ĮĄ▓óį┌50 dų«║¾ĘĆČ©į┌7.09Ī└0.02.▒M╣▄FANį┌50~91 dÅ─ų«Ū░Ą─(329Ī└40) mgĪżL-1Ž┬ĮĄĄĮ┴╦(47Ī└13) mgĪżL-1Ż¼Ą½╩Ū▀@éĆļAČ╬Ą─╚š«aÜŌ┴┐ŲĮŠ∙×ķ(10.8Ī└0.3) LĪżd-1Ż¼ŽÓ▒╚ĘĆČ©ļAČ╬Ą─(14.4Ī└1.1) LĪżd-1ģs£p╔┘┴╦25%.ė╔▒Ē 1┐╔ų¬Ż¼VSĮĄĮŌ┬╩ę▓║═╚š«aÜŌ┴┐ŽÓ╦ŲŻ¼ļSų°ė╬ļx░▒ØŌČ╚Ž┬ĮĄČ°ĮĄĄ═Ż¼▓ó╬┤│÷¼F┼cų«Ū░Ą─蹊┐[19, 20]ŽÓę╗ų┬Ą─ĮY╣¹.į┌Ą┌91 d═Żų╣╠Ē╝ėHClų«║¾Ż¼pH║═FANųØu╗ųÅ═ĄĮģÆ觎¹╗»Ą─│§╩╝ĘČć·Ż¼VSĮĄĮŌ┬╩║═╚š«aÜŌ┴┐ę▓ļSų«╔Ž╔².┐╔ęŖŻ¼į┌▒ŠīŹ“ץ─Ė▀║¼╣╠ģÆ觎¹╗»ŽĄĮyųąŻ¼ĮĄĄ═pHļm╚╗├„’@ĮĄĄ═┴╦FANĄ─ØŌČ╚Ż¼Ą½╩Ū▓óø]ėą▀_ĄĮŅAŲ┌Ą─£p▌pė╬ļx░▒ī”╬ó╔·╬’Ą─ČŠ║”ū„ė├▀MČ°╠ß╔²ŽĄĮy«aÜŌąį─▄Ą──┐Ą─.ŽÓĘ┤Ż¼Å─pHŽ┬ĮĄ║═╗ž╔²Ą─ā╔éĆļAČ╬Ą─öĄō■Č╝┐╔┐┤│÷Ż¼pHĄ─ĮĄĄ═├„’@ė░Ēæ┴╦ŽĄĮyėąÖC╬’Ą─╦«ĮŌ┬╩║═VSĮĄĮŌ┬╩Ż¼▀MČ°ī¦ų┬«a╦ߊ·║═«a╝ū═ķŠ·Ą─Ąū╬’╣®Įo▓╗ūŃŻ¼ūŅĮKę²Ų«aÜŌ┴┐Ą─Ž┬ĮĄ.
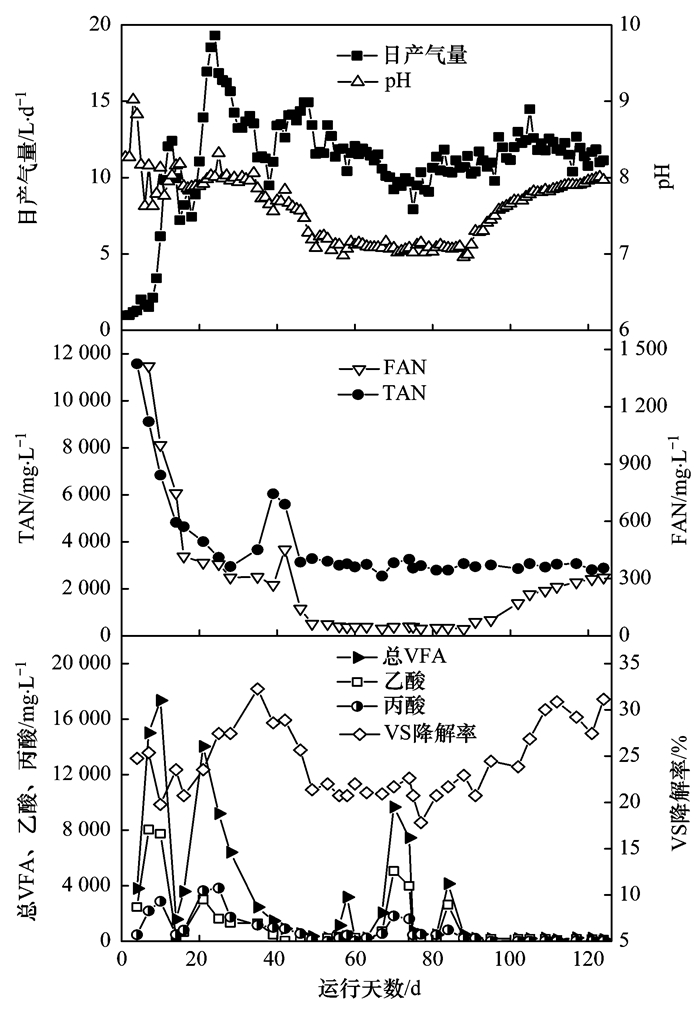
ĪĪĪĪłD 1 R1Ą─ąį─▄ģóöĄļSĢrķgĄ─ūā╗»Ū·ŠĆ
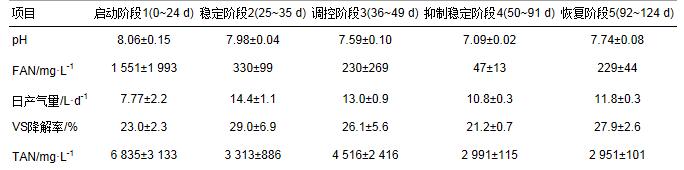
ĪĪĪĪ▒Ē 1 R1į┌▓╗═¼ļAČ╬Ą─Ė„ĒŚųĖś╦(ų├ą┼ģ^ķg95%)
ĪĪĪĪ2.1.2 ╠ßĖ▀ė╬ļx░▒ØŌČ╚ī”«aÜŌ┴┐║═VSĮĄĮŌ┬╩Ą─ė░Ēæ
ĪĪĪĪ┼cR1Ę┤æ¬Ų„▓╗═¼Ż¼R2╩Ū═©▀^╠ßĖ▀TANĄ─ØŌČ╚üĒš{┐žFANĄ─ØŌČ╚.ė╔łD 2┐╔ų¬Ż¼R2į┌åóäėļAČ╬(0~39 d)║═R1ŽÓ╦ŲŻ¼«aÜŌ┴┐▓©äė▌^┤¾Ż¼į┌Ą┌15 d║═Ą┌26 dĘųäe│÷¼F«aÜŌĖ▀ĘÕ. R2į┌40 dų«║¾▀_ĄĮĘĆČ©Ż¼ĘĆČ©ļAČ╬(40~56 d)╚š«aÜŌ┴┐×ķ(12.0Ī└0.3) LĪżd-1Ż¼VSĮĄĮŌ┬╩×ķ(34.1Ī└1.1)%Ż¼╝ū═ķ║¼┴┐╝s×ķ64.8%Ż¼Ė„ĒŚąį─▄ģóöĄ┼cų«Ū░ĘĆČ©▀\ąąĄ─╬█─ÓĖ▀║¼╣╠ģÆ觎¹╗»Ę┤æ¬Ų„ĮėĮ³[21].ė╔łD 2║═▒Ē 2┐╔ų¬Ż¼ļSų°FANĄ─╠ß╔²Ż¼╚š«aÜŌ┴┐║═VSĮĄĮŌ┬╩Č╝│÷¼F┴╦├„’@Ž┬ĮĄŻ¼ŽÓ▒╚ĘĆČ©ļAČ╬Ęųäe£p╔┘┴╦14.2%║═33.7%.ļm╚╗═Ō╝ė░▒Ą¬ų«║¾Ż¼ė╔ė┌NH4Cl┼c╬█─ÓųąĄ─OH-─▄▀MąąĘ┤æ¬Ż¼ī¦ų┬į┌╠ß╔²░▒Ą¬│§Ų┌H+ØŌČ╚į÷╝ėŻ¼pHėą╦∙Ž┬ĮĄ.Ą½╩ŪŻ¼pHĄ─ūā╗»Ę∙Č╚▌^ąĪŻ¼70 dęį║¾ŽĄĮypHėųųØu╗ųÅ═Ż¼▒M╣▄ė╬ļx░▒ØŌČ╚│¼▀^500 mgĪżL-1Ż¼Ą½Ž¹╗»ŽĄĮy╚į╚╗─▄▒Ż│ųĘĆČ©▀\ąąŻ¼šf├„ŽĄĮy╠Äė┌Ī░ęųųŲĄ─ĘĆČ©ĀŅæBĪ▒[3].

ĪĪĪĪłD 2 R2Ą─ąį─▄ģóöĄļSĢrķgĄ─ūā╗»Ū·ŠĆ

ĪĪĪĪ▒Ē 2 R2į┌▓╗═¼ļAČ╬Ą─Ė„ĒŚųĖś╦(ų├ą┼ģ^ķg95%)
ĪĪĪĪ2.2 ė╬ļx░▒š{┐žī”╝ÜŠ·ĘN╚║Ą─ė░Ēæ
ĪĪĪĪ«ö═©▀^ĮĄĄ═pHĄ─ĘĮ╩ĮüĒĮĄĄ═R1ŽĄĮyųąė╬ļx░▒ØŌČ╚ĢrŻ¼╝ÜŠ·ĘN╚║Ą─ūā╗»╚ńłD 3╦∙╩Š.Å─ųą┐╔ų¬Ż¼Candidate_division_WS 6 _norankį┌╗ųÅ═ļAČ╬║¼┴┐├„’@╠ßĖ▀.╚╗Č°Ż¼ęčėąčąŠ┐ųą╔ą╬┤ėø▌d▀@ĘN╝ÜŠ·Ą─Š▀¾w╣”─▄Ż¼ę“Č°ę▓¤oĘ©┼ąöÓŲõį┌ŽĄĮyųą╦∙ŲĄ─ū„ė├.
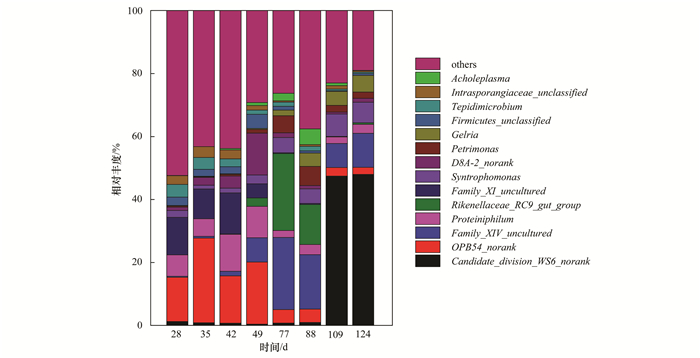
ĪĪĪĪłD 3 R1ųą╝ÜŠ·ŽÓī”žSČ╚ļSĢrķgĄ─ūā╗»
ĪĪĪĪ┴Ēę╗ĘĮ├µŻ¼į┌Ū░ā╔éĆļAČ╬ųąŻ¼ŽÓī”žSČ╚š╝Ū░Äū╬╗Ą─Ęųäe×ķOPB 54 _norank(14%~26%)ĪóProteiniphilum(7%~11.7%)ĪóFamily_XIV_uncultured(5%~13%)║═D8A-2_norank(1%~3.9%Ż¼ŲõųąĄ┌49 dš╝13.2%)Ż¼į┌ęųųŲĘĆČ©ļAČ╬▀@ą®╬ó╔·╬’Ą─ŽÓī”žSČ╚╚½▓┐Č╝Ž┬ĮĄĄĮ5%ęįŽ┬Ż¼’@╚╗ļSų°pHŚl╝■Ą─Ė─ūāŻ¼▀@ą®╝ÜŠ·Ą─╔·ķL║═┤·ųx╩▄ĄĮ┴╦ė░Ēæ. Proteiniphilum╩Ūę╗ĘN«a╦ߊ·Ż¼ų„ꬫa╬’╩Ūęę╦ß║═▒¹╦߯¼─▄ē“▀MąąĮŌļ├ū„ė├ĮĄĮŌĄ░░ū┘|▓ó«a╔·NH3[22].▒ŠīŹ“×ųąŻ¼ŽÓī”žSČ╚ļSpHŽ┬ĮĄĘ∙Č╚ūŅ┤¾Ą─╝ÜŠ·╩ŪOPB54 _norank. HaoĄ╚[23]└¹ė├ęę╦ß×ķ╗∙┘|▀MąąĖ▀£žģÆ觎¹╗»Ż¼▓óįOų├┴╦Ą═TAN (2 600 mgĪżL-1)║═Ė▀TAN (7 000 mgĪżL-1)ā╔ĮMīŹ“ׯ¼░l¼FOPB54(Š▀¾w╣”─▄╬┤ų¬)į┌ā╔ĘN░▒Ą¬ØŌČ╚Ž┬Š∙╩Ū║¼┴┐ūŅČÓĄ─╝ÜŠ·Ż¼Ą½╩Ūį┌Ą═FANŽ┬ŽÓī”žSČ╚Ė³Ė▀Ż¼▀@┼c▒ŠčąŠ┐Ą─ĮY╣¹▓ó▓╗ę╗ų┬.│²┤╦ų«═ŌŻ¼ę╗ą®Ą┌ę╗ļAČ╬║¼┴┐▌^╔┘Ą─╝ÜŠ·╚ńTepidimicrobium(3.9%)ĪóMacellibacteroides(1.8%)ĪóCaldicoprobacter(1.5%)ę▓Č╝ļSų°pHš{┐žė╬ļx░▒Ž┬ĮĄČ°ĘN╚║žSČ╚├„’@ĮĄĄ═. Tepidimicrobium─▄ĘųĮŌŲŽ╠č╠ŪĄ╚ČÓĘN╠ŪŅÉŻ¼ę▓┐╔ęįĮĄĮŌĄ░░ūŅÉĄ─╬’┘|Ż¼«a╬’×ķęę╦ßĪóęę┤╝ĪóH2║═CO2Ą╚Ż¼ėąčąŠ┐▒Ē├„TepidimicrobiumūŅ▀mę╦Ą─pHĘČć·╩Ū8~8.5[24, 25]Ż¼▒ŠīŹ“×ųąpHĮĄĄ═ĄĮ7.1┐╔─▄Ģ■╩ŪęųųŲįōŠ·ī┘╔·ķLĄ─ę╗éĆįŁę“.═¼śėŪķørŽ┬Ż¼ė╔ė┌Macellibacteroides║═Caldicoprobacter╩Ū─▄ĘųĮŌ╠ŪŅÉĄ─╝ÜŠ·Ż¼╦¹éāĄ─ūŅĮK┤·ųx«a╬’ę▓×ķąĪĘųūėĄ─VFAĪóH2║═CO2Ą╚[26, 27]Ż¼▀@ā╔ŅÉ╝ÜŠ·žSČ╚Ą─Ž┬ĮĄę▓į┌ę╗Č©│╠Č╚╔ŽĮŌßī┴╦ėąÖC╬’║¼┴┐╔²Ė▀Ą─įŁę“.ė╔┤╦┐╔ęŖŻ¼═©▀^pHš{┐žė╬ļx░▒ØŌČ╚╩╣Ą├ŽĄĮy└Ē╗»ŁhŠ│▓╗▀mę╦ė┌įŁ▒Šę╗ą®žSČ╚▌^┤¾Ą─ĮĄĮŌ╠ŪŅÉ║═Ą░░ūĄ─«a╦ߊ·Ą─╔·ķLŻ¼Å─Č°╩╣Ą├VSĮĄĮŌ┬╩Ž┬ĮĄ.
ĪĪĪĪė╔łD 3┐╔ų¬Ż¼ļSų°pHĄ─š{┐žŻ¼į┌ęųųŲĘĆČ©ļAČ╬Ż¼└Ē蹊·ī┘(Rikenellaceae_RC9 _gut_group)║═Family_XIV_unculturedĄ─ŽÓī”žSČ╚Å─ų«Ū░Ą─▓╗ūŃ1%Ęųäe╔Ž╔²ų┴24.3%║═22.9%Ż¼Petrimonas║═Acholeplasmaę▓Å─▓╗ūŃ1%Ęųäeį÷ķLĄĮ1.1%║═4.9%.▀@ŲõųąŻ¼└Ē蹊·ī┘Rikenellaceae▓╗āH┐╔ęįĘųĮŌ╠╝╦«╗»║Ž╬’Č°Ūę▀Ć┐╔ęįĮĄĮŌĄ░░ū┘|[28].Č°ųĄĄ├ūóęŌĄ─╩ŪŻ¼ļSų°ė╬ļx░▒ØŌČ╚Ą─╔Ž╔²╗ųÅ═Ż¼▀@ą®╬ó╔·╬’ĘN╚║ŽÓī”öĄ┴┐ėų├„’@Ž┬ĮĄŻ¼▒╚╚ńRikenellaceae_RC9 _gut_groupŽÓī”žSČ╚ėųčĖ╦┘Ž┬ĮĄĄĮ1%ęįŽ┬Ż¼Family_XIV_unculturedę▓ĮĄĄ═ų┴2.8%.ėąčąŠ┐▒Ē├„╔Ž╩÷▀@ą®Š·ī┘ļm╚╗ę▓Č╝╩Ū▀Mąą╠ŪŅÉĮĄĮŌĄ─«a╦ߊ·[29, 30]Ż¼Ą½Å─ĮY╣¹üĒ┐┤Ųõ┤·ųx─▄┴”┼cTepidimicrobiumĪóMacellibacteroides║═CaldicoprobacterŽÓ▒╚├„’@▌^╚§.╗źĀIå╬░¹Š·ī┘SyntrophomonasĄ─ĘN╚║žSČ╚į┌ęųųŲĘĆČ©ļAČ╬ę▓ėą├„’@į÷ķLŻ¼Ą½į┌╗ųÅ═ļAČ╬▓óø]ėąŽ┬ĮĄŻ¼▒Ē├„┴╦▀@ŅÉ╬ó╔·╬’┴╝║├Ą─pH▀mæ¬ĘČć·.╗źĀIå╬░¹Š·ī┘Syntrophomonas╩Ūę╗ĘN╗źBą═Ą─Š▀ėąč§╗»ę╗ą®ųžę¬Ą─ųąķg«a╬’╚ń▒¹╦ß║═ČĪ╦ߥ─╣”─▄Ą─╝ÜŠ·Ż¼┤·ųx«a╬’×ķęę╦ß║═H2Ż¼įō╝ÜŠ·▒žĒÜę¬║═─▄ē“Ž¹║─H2Ą─╬ó╔·╬’╣▓═¼┼ÓB▓┼─▄ē“╔·ķL[31]Ż¼ī┘ė┌╗źBą═Ą─ĮĄĮŌ═ŠÅĮ. Syntrophomonasį┌FAN╔²Ė▀ĢrĢ■╩▄ĄĮęųųŲŻ¼į┌蹊┐▓╗═¼Ą─╣ż│╠ęÄ─Ż[32]╗“š▀īŹ“×╩ęęÄ─Ż[33]Ą─ģÆč§Ę┤æ¬Ų„ųąĄ─╬ó╔·╬’ĘN╚║ĮYśŗĢr░l¼FĖ▀ė╬ļx░▒ØŌČ╚Ą─Ę┤æ¬Ų„└’└²╚ńSyntrophomonasĪóPelotomaculumĪóDesulfobulbusĄ╚╗źBą═╬ó╔·╬’▌^Ą═ė╬ļx░▒Ą─Ę┤æ¬Ų„║¼┴┐Ė³╔┘Ż¼’@╩Š╗źBą═┤·ųxĖ³ęū╩▄ĄĮė╬ļx░▒Ą─ęųųŲ.«ö×ķ┴╦ĮĄĄ═ė╬ļx░▒ØŌČ╚Č°┐žųŲpHųĄų┴7.0ų«║¾Ż¼ŽĄĮyųą│ąō·╠ŪŅÉ║═Ą░░ūĄ─ĮĄĮŌĄ─╬ó╔·╬’ŅÉą═░l╔·┴╦▐D╗»Ż¼ų«Ū░žSČ╚▌^┤¾Ą─įōŅÉ╝ÜŠ·ĘN╚║ŽÓī”öĄ┴┐Č╝│÷¼FĮĄĄ═Ż¼Č°┴Ēę╗ą®ų«Ū░žSČ╚▌^ąĪĄ─╝ÜŠ·║¼┴┐į÷╝ėŻ¼┼c┤╦═¼ĢrĘ┤æ¬Ų„Ą─ėąÖC╬’ĮĄĮŌąį─▄║═«aÜŌąį─▄│÷¼FŽ┬ĮĄ.
ĪĪĪĪČ°«ö═©▀^═Ō╝ė░▒Ą¬╠ßĖ▀R2ŽĄĮyųąė╬ļx░▒ØŌČ╚ĢrŻ¼╝ÜŠ·ĘN╚║Ą─ūā╗»╚ńłD 4╦∙╩Š, R2Ę┤æ¬Ų„ųą3ĘNų„ꬥ─ā×ä▌╝ÜŠ·ĘN╚║Ż¼į┌åóäėļAČ╬Ż¼Tepidimicrobium║═ProteiniborusĄ─ŽÓī”žSČ╚Ęųäeį┌Ą┌21 d║═Ą┌16 d▀_ĄĮĖ„ūįĄ─ūŅĖ▀ųĄ33.8%║═15.8%.Č°║¾Ż¼ā╔ĘN╝ÜŠ·Č╝į┌35 dŽ┬ĮĄŻ¼Ęųäe×ķ18.3%║═6.2%.╝ÜŠ·ĘN╚║Ą─žSČ╚į┌ĘĆČ©ļAČ╬▒Ż│ų╗∙▒Š▓╗ūāŻ¼│²┴╦Anaerobrancaį┌Ą┌42 dėąę╗éĆ╝żį÷.«öĄ┌56 dķ_╩╝Ż¼ļSų°R2ā╚ė╬ļx░▒ØŌČ╚Ą─▓╗öÓ╔Ž╔²Ż¼Anaerobrancaę▓│÷¼F┴╦├„’@Ą─ļSų«╔Ž╔²Ą─┌ģä▌Ż¼Č°Tepidimicrobium║═Proteiniborusät▒Ē¼F│÷ļSų°FAN╔Ž╔²Č°ĮĄĄ═Ą─┌ģä▌. Tepidimicrobium╩Ūę╗ĘNĮŌļ╦ą═Ą─╝ÜŠ·Ż¼┐╔ęįęįČÓĘNĄ░░ū┘|Ą─╬’┘|×ķ╗∙┘|üĒŠS│ųūį╝║Ą─╔·ķL[25]Ż¼į┌R1║═R2ā╔ĮMīŹ“×ųąŻ¼Č╝░l¼F┴╦TepidimicrobiumĄ─ŽÓī”žSČ╚ļSų°ė╬ļx░▒Ą─ūā╗»├„’@ĄžĖ─ūāŻ¼╝┤▓╗šō╩Ūė╬ļx░▒ØŌČ╚╠ßĖ▀▀Ć╩ŪĮĄĄ═Ż¼TepidimicrobiumĄ─žSČ╚Č╝ėą╦∙Ž┬ĮĄ.║═TepidimicrobiumŅÉ╦ŲŻ¼Proteiniborusę▓┐╔ęįį┌pHųĄ6~8Ą─ĘČć·└’└¹ė├Ą░░ū┘|ū„×ķ░lĮ═Ąū╬’[34]. Anaerobranca▒Š┘|╔Žę▓╩ŪĘųĮŌĄ░░ū┘|Ą─╝ÜŠ·[35]. ProweĄ╚[36]░l¼FAnaerobrancaų╗░³║¼ÄūĘN┐╔ęįģÆč§░lĮ═Ą░░ū▐D╗»×ķęę╦ߥ─ģÆč§╬ó╔·╬’.▒ŠīŹ“×ųąĄ┌42 d AnaerobrancaĄ─į÷ČÓ═¼Ģrę▓░ķļSų°ė╬ļx░▒ØŌČ╚Ą─╔Ž╔²Ż¼┐╔ęį┐┤│÷«öė╬ļx░▒ØŌČ╚ūāĖ▀Ą─Ģr║“Ż¼Anaerobrancaį┌š¹éĆĄ░░ūĄ─ĘųĮŌ▀^│╠ųą╦∙ŲĄ─ū„ė├Ė³┤¾.Č°Anaerobrancaī”Ą░░ū┘|Ą─ĮĄĮŌ╦┘┬╩┐╔─▄ąĪė┌Tepidimicrobium║═ProteiniborusŻ¼Å─Č°ī¦ų┬ŽĄĮyVSĮĄĮŌ┬╩Ą─Ž┬ĮĄ.
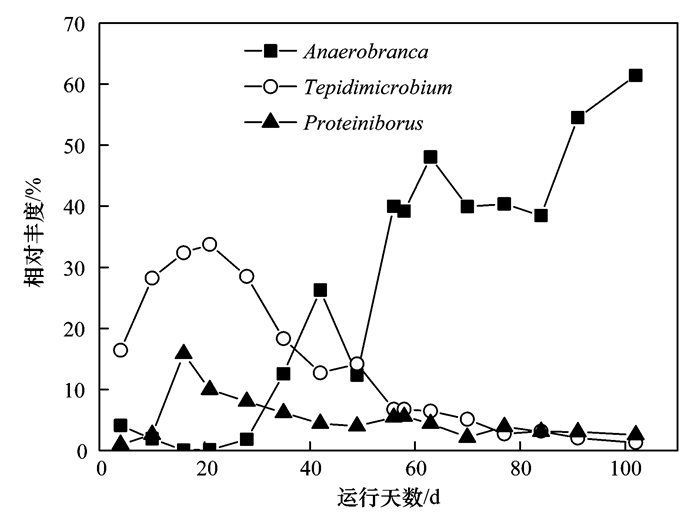
ĪĪĪĪłD 4 R2ųą3ĘNā×ä▌╝ÜŠ·ŽÓī”žSČ╚ļSĢrķgĄ─ūā╗»
ĪĪĪĪ2.3 ė╬ļx░▒š{┐žī”«a╝ū═ķŠ·ĘN╚║Ą─ė░Ēæ
ĪĪĪĪė╔łD 5┐╔ų¬Ż¼R1Ę┤æ¬Ų„ųąė╬ļx░▒ØŌČ╚Ž┬ĮĄų«║¾Ż¼Üõ└¹ė├ą═Ą─╝ū═ķ─ꊷī┘(Methanoculleus)Ą─ŽÓī”žSČ╚├„’@ĮĄĄ═Ż¼Å─13%Ž┬ĮĄĄĮ1.9%Ż¼▓óŪęį┌╗ųÅ═ļAČ╬ę▓╬┤─▄═Ļ╚½╗ųÅ═ų┴įŁėą╦«ŲĮŻ¼└^└mŽ┬ĮĄų┴▓╗ūŃ1%.Č°š¹éĆ▀^│╠ųąŻ¼ęę╦ßą═Ą─╝ū═ķ░╦»BŪ“Š·ī┘(Methanosarcina)ŽÓī”žSČ╚ę╗ų▒┤¾ė┌80%Ż¼į┌┼c╝ū═ķ─ꊷī┘Ą─ĖéĀÄųąŻ¼ļSų°ė╬ļx░▒ØŌČ╚║═pHĄ─Ž┬ĮĄŻ¼ųØuš╝ō■┴╦Ė³┤¾Ą─ā×ä▌Ż¼ūŅ║¾žSČ╚╔§ų┴▀_ĄĮ97%.Ą½╩Ūę“×ķŽĄĮyĄ─«aÜŌ┴┐║═VSĮĄĮŌ┬╩ļSų°FAN║═pHĄ─Ž┬ĮĄČ°ĮĄĄ═Ż¼×ķ┴╦Ė³£╩┤_Ąžė^▓ņė╬ļx░▒╝░pHī”«a╝ū═ķŠ·ĘN╚║ė╚Ųõ╩Ū╝ū═ķ░╦»BŪ“Š·(Methanosarcina)Ą─ė░ĒæŻ¼═©▀^Č©┴┐PCRĄ─Ęų╬÷Č°½@Ą├Ųõ16S rRNA╗∙ę“ØŌČ╚Ą─ūā╗»┌ģä▌üĒČ©┴┐Ęų╬÷╝ū═ķŠ·ĘN╚║Ą─äėæBūā╗».ė╔łD 6ų¬Ż¼R1Ę┤æ¬Ų„ā╚ŽÓī”žSČ╚ūŅ┤¾Ą─╝ū═ķ░╦»BŪ“Š·Ą─16S rRNA╗∙ę“ØŌČ╚(ęįر─ÓėŗŻ¼Ž┬═¼)į┌Ą┌28 d║═35 dĘųäe×ķ(1.27Ī┴106Ī└8.0Ī┴104) copiesĪżg-1║═(1.19Ī┴106Ī└9.89Ī┴104) copiesĪżg-1Ż¼ų«║¾▒M╣▄pH▓╗öÓĮĄĄ═Ż¼Ą½ę└╚╗ŠS│ųį┌═¼ę╗éĆöĄ┴┐╝ēā╚Ż¼▓©äėūā╗»▓╗┤¾.Č°ĄĮ┴╦Ą┌88 d 16S rRNA╗∙ę“ØŌČ╚├„’@Ž┬ĮĄŻ¼×ķ(1.27Ī┴105Ī└1.22Ī┴104) copiesĪżg-1Ż¼▀@ĢrŽĄĮypHųĄęčĮøį┌7.1ŠS│ų┴╦38 dŻ¼╝s2éĆSRTŻ¼’@╚╗ų«Ū░pH×ķ7ū¾ėęĄ─ķLĢrķgū„ė├ęčĮø╩╣Ą├▌^ČÓĄ─╝ū═ķ░╦»BŪ“Š·╩▄ĄĮĄū╬’│ų└m╣®Įo▓╗ūŃĄ─ė░Ēæ.ų«║¾ļSų°FAN║═pHĄ─╗ž╔²Ż¼╝ū═ķ░╦»BŪ“Š·16S rRNA╗∙ę“ØŌČ╚ėų╗ž╔²ų┴106 copiesĪżg-1Ą─öĄ┴┐╝ē╦«ŲĮ.┐╔ęŖ«a╝ū═ķŠ·ĘN╚║į┌═©▀^pHš{┐žė╬ļx░▒Ą─▀^│╠ųąė╔ė┌╝ÜŠ·ĘN╚║ĮYśŗĄ─ė░ĒæČ°╩▄ĄĮ╗∙┘|╣®Įo▓╗ūŃĄ─ęųųŲŻ¼Ą½ļSų°pHĄ─╗ž╔²ėų▌^┐ņ╗ųÅ═┴╦╗Ņąį.
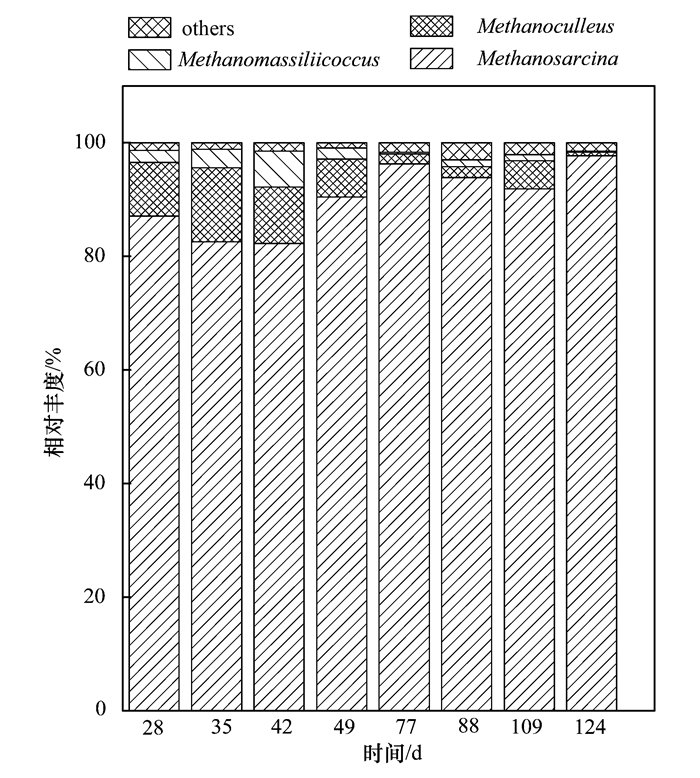
ĪĪĪĪłD 5 R1Ą─╣┼Š·į┌ī┘Ą─╦«ŲĮ╔ŽļSĢrķgĄ─ūā╗»
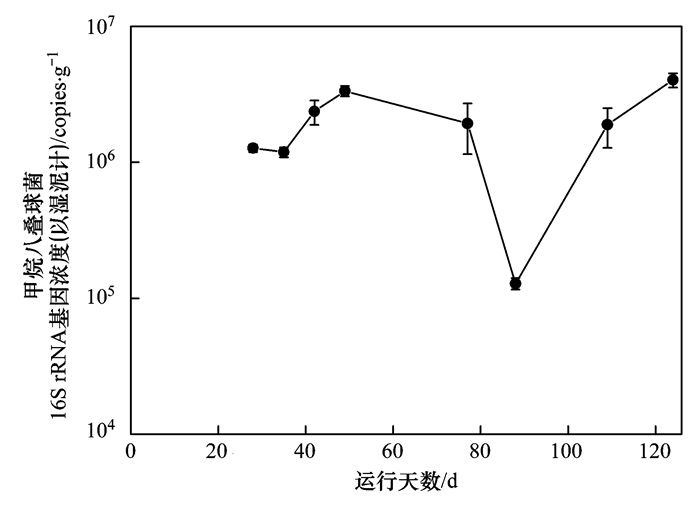
ĪĪĪĪłD 6 R1╝ū═ķ░╦»BŪ“Š·öĄ┴┐ļSĢrķgĄ─ūā╗»(n=3)
ĪĪĪĪR2śėŲĘĄ─╣┼Š·ČÓśėąįĘų╬÷ĮY╣¹╚ńłD 7╦∙╩ŠŻ¼ęę╦ßą═Ą─╝ū═ķ░╦»BŪ“Š·Methanosarcina×ķā×ä▌ĘN╚║Ż¼į┌š¹éĆīŹ“×▀^│╠ųąš╝╣┼Š·Ą─▒╚└²ę╗ų▒į┌92%ęį╔ŽŻ¼šf├„╝ū═ķ░╦»BŪ“Š·┐╔─▄▓óø]ėą╩▄ĄĮė╬ļx░▒ØŌČ╚╠ßĖ▀Ą─ęųųŲ.Č©┴┐PCRĄ─ĮY╣¹ę▓▒Ē├„į┌ė╬ļx░▒ØŌČ╚Å─(400Ī└173) mgĪżL-1╠ßĖ▀ĄĮ(526Ī└25) mgĪżL-1ų«║¾Ż¼R2ā╚Ą─╝ū═ķ░╦»BŪ“Š·Ą─16S rRNA╗∙ę“ŲĮŠ∙ØŌČ╚Å─(3.43Ī┴106Ī└2.91Ī┴106) copiesĪżg-1ūā╗»×ķ(3.58Ī┴106Ī└1.18Ī┴106) copiesĪżg-1Ż¼öĄ┴┐╗∙▒ŠŽÓ═¼.ę“Č°Ż¼ėąÖC╬’ĮĄĮŌ╩▄ĄĮęųųŲĄ─ų„ę¬įŁę“æ¬įōüĒį┤ė┌╝ÜŠ·ĘN╚║ĮYśŗ╦∙╩▄Ą─ė░Ēæ[37].Š▀¾wģóęŖ╬█╦«īÜ╔╠│Ū┘Y┴Ž╗“http://www.jianfeilema.cnĖ³ČÓŽÓĻP╝╝ąg╬─ÖnĪŻ
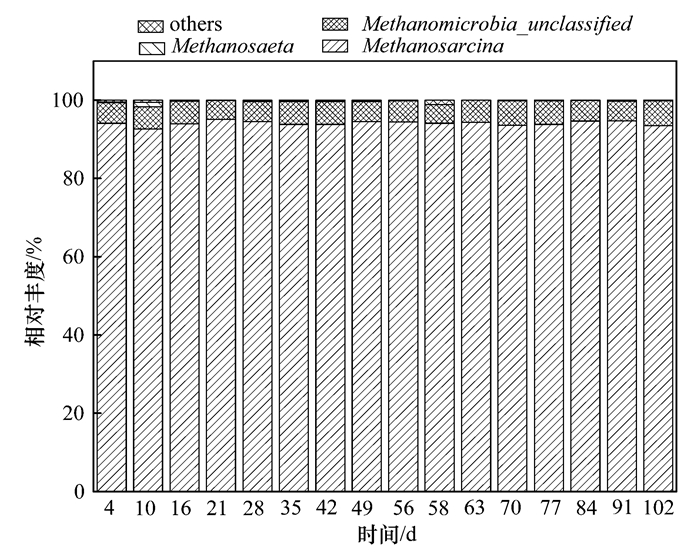
ĪĪĪĪłD 7 R2Ą─╣┼Š·į┌ī┘Ą─╦«ŲĮ╔ŽļSĢrķgĄ─ūā╗»
ĪĪĪĪ3 ĮYšō
ĪĪĪĪ(1)«öFANÅ─(400Ī└173) mgĪżL-1╠ßĖ▀ĄĮ(526Ī└25) mgĪżL-1║¾Ż¼ŽÓ▒╚š²│ŻĄ─Ž¹╗»ĀŅæBŻ¼ŽĄĮyĄ─╚š«aÜŌ┴┐║═VSĮĄĮŌ┬╩ĘųäeĮĄĄ═┴╦14.2%║═33.7%Ż¼Ą½ę└╚╗─▄į┌ęųųŲĄ─ĀŅæBŽ┬ĘĆČ©▀\ąą.
ĪĪĪĪ(2)«ö═©▀^pHš{┐ž╩╣Ą├FANÅ─(330Ī└99) mgĪżL-1Ž┬ĮĄų┴(47Ī└13) mgĪżL-1Ż¼ŽĄĮyĄ─╚š«aÜŌ┴┐║═VSĮĄĮŌ┬╩ĘųäeĮĄĄ═┴╦25%║═26.9%Ż¼Č°ļSų°pH║═FAN╗ųÅ═ų┴š²│Ż╦«ŲĮŻ¼ŽĄĮyĄ─«aÜŌąį─▄║═VSĮĄĮŌ┬╩ėų╗ųÅ═ų┴įŁėą╦«ŲĮŻ¼ŽÓ▒╚ęųųŲĘĆČ©ļAČ╬Ęųäe╠ßĖ▀┴╦9.3%║═31.6%.
ĪĪĪĪ(3)▒Š╬─ųąĄ─ā╔ĘNė╬ļx░▒š{┐žĘĮ╩ĮČ╝Ģ■ė░Ēæ╬█─ÓĖ▀║¼╣╠ģÆ觎¹╗»ŽĄĮyųą╝ÜŠ·Ą─ĘN╚║ĮYśŗ.═Č╝ė═ŌüĒ░▒Ą¬║¾Ż¼▀^Ė▀Ą─FAN (500~600 mgĪżL-1)Ģ■╩╣Ą├ĮĄĮŌĄ░░ūĄ─╝ÜŠ·ĘNŅÉ│»ų°Ą░░ūĮĄĮŌ┬╩Ė³Ą═Ą─╝ÜŠ·Š·╚║▐DūāŻ¼Ą½▓óø]ėąė░ĒæĄĮų„ꬥ─«a╝ū═ķŠ·(╝ū═ķ░╦»BŪ“Š·)Ą─╔·ķL;┴Ēę╗ĘĮ├µŻ¼═©▀^ĮĄĄ═pHüĒĮĄĄ═ė╬ļx░▒Ą─ØŌČ╚▓ó▓╗ę╗Č©─▄╩╣Ą├ąį─▄╠ßĖ▀Ż¼Ę┤Č°┐╔─▄Ģ■ė╔ė┌pHĄ─ūā╗»ė░Ēæę╗ą®Ą─╠ŪŅÉ║═Ą░░ū┘|ŅÉ░lĮ═╝ÜŠ·Ą─╗ŅąįŻ¼ī¦ų┬╦«ĮŌą¦┬╩Ą─Ž┬ĮĄŻ¼ę²░l║¾└m«a╝ū═ķŠ·Ą─Ąū╬’╣®Įo▓╗ūŃŻ¼Ž„╚§ŽĄĮy«aÜŌąį─▄.(üĒį┤╝░ū„š▀Ż║═¼Ø·┤¾īWŁhŠ│┐ŲīW┼c╣ż│╠īWį║ ┤„Ģį╗óĪó║╬▀MĪóć└║«Īó└ŅīÄĪóČĪį┬┴ßĪóČŁ×IĪó┤„¶ß¶ß)

