Ė▀░▒Ą¬╣żśIÅU╦«╠Ä└Ē╣ż╦ć蹊┐
ųąć°╬█╦«╠Ä└Ē╣ż│╠ŠW ĢrķgŻ║2017-5-15 8:50:42
╬█╦«╠Ä└Ē╝╝ąg | ģRŠ█╚½Ū“Łh▒Ż┴”┴┐Ż¼ĮĄĄ═Ų¾śIų╬╬█│╔▒Š
ĪĪĪĪ20╩└╝o90─Ļ┤·Ż¼║╔╠mĄ─MulderĄ╚į┌╠Ä└Ē╣żśIÅU╦«Ą─Ę┤Ž§╗»┴„╗»┤▓ųą░l¼F┴╦ę╗ĘNą┬ą═╝ÜŠ·Ī¬Ī¬ģÆč§░▒č§╗»╝ÜŠ· (anaerobic ammonia oxidation bacteriaŻ¼AnAOB).įōŅÉ╝ÜŠ·┐╔ęįį┌╚▒觌l╝■Ž┬ęįü厧╦ß¹}×ķļŖūė╩▄¾wīó░▒Ą¬▐D╗»×ķĄ¬ÜŌ. 1999─ĻŻ¼StrousĄ╚▓╔ė├├▄Č╚╠▌Č╚ļxą─Ę©│╔╣”Ęųļx┴╦AnAOBŠ·ųĻŻ¼▓ó═©▀^16S rRNA╗∙ę“£yą“Ż¼┤_šJ┴╦AnAOB╩ŪĖĪ├╣ĀŅŠ·Ą─│╔åT.
ĪĪĪĪ┼cé„ĮyĄ─Ž§╗»Ę┤Ž§╗»╣ż╦ćŽÓ▒╚Ż¼ģÆč§░▒č§╗» (anaerobic ammonia oxidationŻ¼ANAMMOX) ╣ż╦ćŠ▀ėą¤oąĶ═Ō╝ė╠╝į┤║═╣®č§äė┴”Ž¹║─Ż¼Ę┤æ¬▀^│╠ųąCO2┼┼Ę┼┴┐╔┘║═╩ŻėÓ╬█─Ó«a┴┐╔┘Ą╚ā׳c.─┐Ū░įō╣ż╦ćęčĮøė├ė┌Ė▀░▒Ą¬╣żśIÅU╦«Ą─╠Ä└ĒųąŻ¼╚ń╬█─ÓŽ¹╗»ę║Īó└¼╗°ØB×Vę║║═Bžił÷ÅU╦«Ą╚.
ĪĪĪĪģÆč§░▒č§╗»╣ż╦ćļm╚╗ėąųTČÓā׳cŻ¼Ą½AnAOB╔·ķLŠÅ┬²Ż¼▒Čį÷Ģrķg×ķ7~22 dŻ¼ī¦ų┬ANAMMOXĘ┤æ¬Ų„åóäėĢrķgķLŻ¼╩ŪŽ▐ųŲģÆč§░▒č§╗»╣ż╦ć┤¾ęÄ─Żæ¬ė├Ą─ūŅų„ę¬ę“╦ž.ę“┤╦Ż¼▀xō±║Ž▀mĄ─Ę┤æ¬Ų„Ż¼£p╔┘AnAOBĄ─┴„╩¦Ż¼╩Ū│╔╣”æ¬ė├ģÆč§░▒č§╗»╣ż╦ćĄ─ĻPµI.─┐Ū░Ż¼ė├ė┌ģÆč§░▒č§╗»╝╝ągĄ─Ę┤æ¬Ų„ėą╔Ž┴„╩Į╬█─Ó┤▓Ę┤æ¬Ų„ (USB)Īó┼·ą“╩Įķgą¬Ę┤æ¬Ų„ (SBR)║═ģÆ觚█┴„░ÕĘ┤æ¬Ų„ (ABR)Ą╚.▒ŠīŹ“×▀xō±╔Ž┴„╩ĮģÆč§▀^×V┤▓Ę┤æ¬Ų„ (upflow blanket filter, UBF)Ż¼UBFĘ┤æ¬Ų„╩Ūė╔ (╔Ž┴„╩Į╬█─Ó┤▓) USB║═ģÆč§▀^×VŲ„ (AF) śŗ│╔Ą─Å═║Ž╩ĮĘ┤æ¬Ų„Ż¼Š▀ėą┴╝║├Ą─╔·╬’Įž┴¶─▄┴”║═ęūą╬│╔Ņw┴Ż╬█─ÓĄ╚╠ž³c.
ĪĪĪĪAnAOBų┴Į±╚į╬┤½@Ą├╚╦╣żĄ─╝ā┼ÓBŻ¼Č°Ė▀═©┴┐£yą“╝╝ągĄ─░lš╣×ķĶbČ©ŁhŠ│ųąĄ─╬ó╔·╬’╚║┬õĄ─ČÓśėąį╠ß╣®┴╦ėą┴”Ą─╣żŠ▀.▒ŠčąŠ┐▓╔ė├Illumina Hisep PE250£yą“ŲĮ┼_ī”UBFģÆč§░▒č§╗»Ę┤æ¬Ų„ā╚Ą─╬ó╔·╬’▀MąąĖ▀═©┴┐£yą“Ż¼Ęų╬÷┴╦ANAMMOXĘ┤æ¬Ų„ā╚╬ó╔·╬’Ą─Ęų▓╝ŪķørŻ¼ęįŲ┌×ķANAMMOXĘ┤æ¬Ų„ā╚╬ó╔·╬’╚║┬õŽÓī”žSČ╚╝░╚║┬õĮYśŗūā╗»╠ß╣®└Ēšōę└ō■.
ĪĪĪĪ1 ▓─┴Ž┼cĘĮĘ©1.1 īŹ“×čbų├
ĪĪĪĪĘ┤æ¬Ų„×ķUBFģÆč§░▒č§╗»Ę┤æ¬Ų„Ż¼ė╔ėąÖC▓Ż┴¦ųŲ│╔Ż¼│╩łAų∙ą╬Ż¼┐é╚▌Ęe×ķ3.94 LŻ¼ėąą¦╚▌Ęe×ķ2.1 LŻ¼ų▒ÅĮ×ķ8 cmŻ¼┐éĖ▀Č╚×ķ66 cmŻ¼ā╚ų├20 cmĖ▀Ą─╠Ņ┴ŽīėŻ¼═Ō╣³║┌▓╝Ż¼▒▄├Ō╣Ōī”ģÆč§░▒č§╗»Š·Ą─ęųųŲ.ÅU╦«═©▀^╚õäė▒├Å─Ę┤æ¬Ų„Ą─Ąū▓┐▀B└m▒├╚ļŻ¼┼c╬█─Ó╗ņ║Ž║¾Ż¼Ę┤æ¬╔·│╔Ą─ÜŌ¾wÅ─Ę┤æ¬Ų„Ēö▓┐Ą─┼┼ÜŌ┐ū┼┼│÷Ż¼│÷╦«ė╔Ę┤æ¬Ų„╔Ž▓┐Ą─ęń┴„č▀┴„│÷.▀M╦«pH┐žųŲį┌7.1~7.8Ż¼į┌│Ż£žŚl╝■Ž┬åóäė.īŹ“×čbų├╚ńłD 1╦∙╩Š.
ĪĪĪĪ
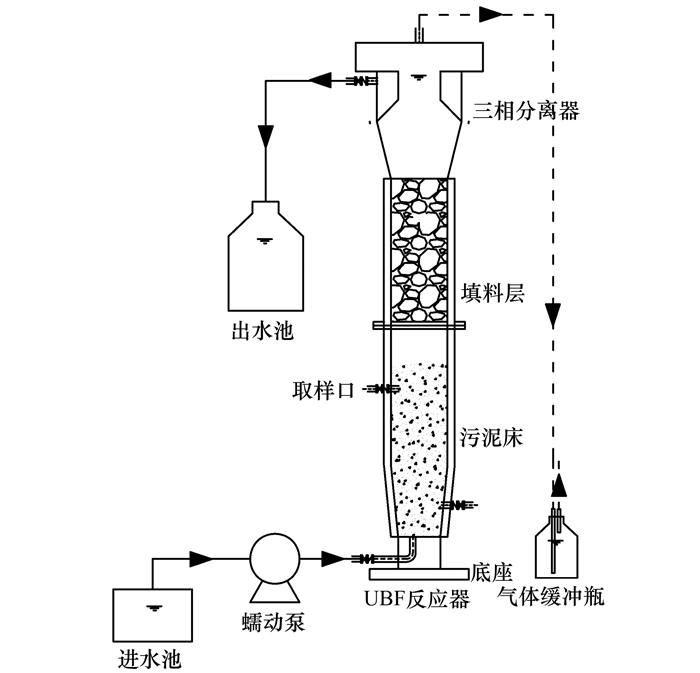
łD 1 īŹ“×čbų├╩ŠęŌ
ĪĪĪĪ1.2 ĮėĘN╬█─Ó║═īŹ“×ė├╦«
ĪĪĪĪĘ┤æ¬Ų„ĮėĘN╬█─ÓüĒūįÅVų▌╩ą×r£“╬█╦«╠Ä└ĒÅS╚▒č§│žŻ¼ĮėĘN┴┐×ķ800 mL.
ĪĪĪĪīŹ“×▓╔ė├─ŻöMÅU╦«Ż¼ų„ę¬│╔Ęų×ķNH4Cl║═NaNO2Ż¼ŲõØŌČ╚░┤īŹ“׹Ķę¬╠Ē╝ė.Ųõ╦³│╔Ęų×ķŻ║NaHCO3 1 000 mg ĪżL-1Ż¼MgSO4 Īż7H2O 473 mg ĪżL-1Ż¼CaCl2 Īż2H2O 180 mg ĪżL-1Ż¼KH2PO4 27 mg ĪżL-1Ż¼╬ó┴┐į¬╦žó±1 mg ĪżL-1║═╬ó┴┐į¬╦žó“ 1 mg ĪżL-1.Ųõųą╬ó┴┐į¬╦žó±×ķŻ║EDTA 5 000 mg ĪżL-1║═FeSO4 Īż7H2O 5 000 mg ĪżL-1.╬ó┴┐į¬╦žó“×ķŻ║ZnSO4 Īż7H2O 430 mg ĪżL-1Ż¼GuSO5 Īż5H2O 240 mg ĪżL-1Ż¼MnCl2 Īż4H2O 990 mg ĪżL-1Ż¼NiCl2 Īż6H2O 190 mg ĪżL-1Ż¼CoCl2Īż6H2O 24 mg ĪżL-1.▀M╦«pHųĄė├¹}╦ßš{╣Ø×ķ7.1~7.8.
ĪĪĪĪ1.3 ╦«śėÖz£yĘĮĘ©
ĪĪĪĪ░▒Ą¬Ż║╝{╩Žįćä®Ęų╣Ō╣ŌČ╚Ę©;ü厧Ą¬Ż║N-(1-▌┴╗∙)-ęęČ■░Ę╣ŌČ╚Ę©;Ž§Ą¬Ż║ūŽ═ŌĘų╣Ō╣ŌČ╚Ę©;┐饬×ķ░▒Ą¬Īóü厧Ą¬║═Ž§Ą¬ų«║═;pHųĄ▓╔ė├pHėŗ£yČ©.
ĪĪĪĪ1.4 ╬ó╔·╬’╚║┬õČÓśėąįÖz£yĘĮĘ©1.4.1 DNA╠ß╚Ī
ĪĪĪĪĘ┤æ¬Ų„▀\ąąų┴Ą┌54 dĢr╚ĪŽ┬īė╬█─Ó20 mL (ś╦ėø×ķCYR110Ż¼╬─ųą▒Ē╩ŠĖ╗╝»ļAČ╬)Ż¼┤╦Ģr╬█─ÓĖ▀Č╚×ķ19.6 cm.Ą┌83 dĢrĘųäe╚Ī╔Žīė╬█─Ó (ś╦ėø×ķCYR111) ║═Ž┬īė╬█─Ó (ś╦ėø×ķCYR112Ż¼╬─ųą▒Ē╩ŠĘĆČ©▀\ąąļAČ╬) Ė„20 mLŻ¼┤╦Ģr╬█─ÓĖ▀Č╚×ķ24.4 cm.Ž┬īė╬█─ÓĄ─╚Īśė┐┌ŠÓļxĄū▓┐4.4 cmŻ¼╔Žīė╬█─ÓĄ─╚Īśė┐┌ŠÓļxĄū▓┐20.7 cm.▓╔ė├Powersoil═┴╚└DNA╠ß╚Īįćä®║ą╠ß╚Ī╗∙ę“ĮMDNA.╠ß╚ĪĄ─DNA╩╣ė├Łéų¼╠Ū─²─zļŖėŠÖz£yDNA╝āČ╚Ż¼╩╣ė├NanodropÖz£yDNAØŌČ╚.
ĪĪĪĪ1.4.2 PCRöUį÷
ĪĪĪĪ▓╔ė├16S rRNA V4ģ^═©ė├ę²╬’Ż¼ę²╬’ą“┴ą×ķ515F 5Īõ-GTGCCAGCMGCCGCGGTAA-3Īõ║═806R 5Īõ-GGACTACHVGGGTWTCTAAT-3ĪõŻ¼öUį÷Ų¼Č╬┤¾ąĪ×ķ300bpū¾ėę.▓╔ė├PCRāxī”╝ÜŠ·16S rRNA╗∙ę“▀MąąPCRöUį÷Ż¼öUį÷Ę┤款wŽĄ (60 ”╠L) ×ķ10x Ex Taq Buffer 6 ”╠LŻ¼dNTP 6 ”╠LŻ¼BSA 0.6 ”╠LŻ¼Ex Taq 0.3 ”╠LŻ¼Primer F 1.2 ”╠LŻ¼Primer R 1.2 ”╠LŻ¼DNA 1 ”╠LŻ¼ddH2O 43.7 ”╠L.░┤ššPCRĘ┤æ¬Śl╝■ (94Īµ 5 minŻ¼94Īµ 30 sŻ¼52Īµ 30 sŻ¼72Īµ 45 sŻ¼72Īµ 10 min) ▀MąąŻ¼31 Cycles PCR═Ļ│╔║¾└¹ė├Łéų¼╠Ū─²─zļŖėŠÖz£yöUį÷«a╬’.
ĪĪĪĪ1.4.3 IlluminaĖ▀═©┴┐£yą“
ĪĪĪĪ▓╔ė├Illumina Hisep PE250£yą“ŲĮ┼_ī”▓╔╝»Ą─śėŲĘ▀MąąĖ▀═©┴┐£yą“.
ĪĪĪĪ1.4.4 öĄō■╠Ä└ĒĘų╬÷ĘĮĘ©
ĪĪĪĪ▓╔ė├Origin 7.5║═AutoCAD 2007▄ø╝■└LųŲłDŻ¼▓ó▓╔ė├SPSS 19.0(IBM IncŻ¼USA) ▄ø╝■ī”öĄō■▀MąąĘų╬÷.
ĪĪĪĪ2 ĮY╣¹┼cėæšō2.1 Ę┤æ¬Ų„Ą─├ōĄ¬ąį─▄
ĪĪĪĪĘ┤æ¬Ų„▀\ąąų┴Ą┌54dĢrŻ¼▒Ż│ųHRT×ķ8h▓╗ūāŻ¼ęį╠ßĖ▀▀M╦«NH4+-N║═NO2--NØŌČ╚Ą─ĘĮ╩Įį÷┤¾Ę┤æ¬Ų„Ą─╚▌Ęežō║╔Ż¼┤╦ĢrNH4+-NĪóNO2--N║═TNĄ─╚ź│²┬╩Ęųäe×ķ76.18%Īó53.47%║═55.66%Ż¼┐饬╚▌Ęežō║╔║═╚ź│²žō║╔Ęųäe×ķ0.56 kg Īż(m3 Īżd)-1║═0.29 kg Īż(m3 Īżd)-1.«öĘ┤æ¬Ų„▀\ąąų┴Ą┌83 dĢrŻ¼NH4+-NĪóNO2--N║═TNĄ─╚ź│²┬╩Ęųäe×ķ99.4%Īó93.1%║═82.8%Ż¼┐饬╚▌Ęežō║╔║═╚ź│²žō║╔Ęųäe×ķ1.25 kg Īż(m3 Īżd)-1║═1.04 kg Īż(m3 Īżd)-1Ż¼šf├„Ę┤æ¬Ų„åóäė│╔╣”.ė╔łD 2┐╔ęį┐┤│÷Ż¼NH4+-NĪóNO2--NØŌČ╚║═NLR│╩╠▌Č╚╔²Ė▀.«öĘ┤æ¬Ų„Ą─┐饬╚▌Ęežō║╔ė╔0.52 kg Īż(m3 Īżd)-1į÷┤¾ų┴2.75 kg Īż(m3 Īżd)-1ĢrŻ¼TNĄ─╚ź│²┬╩ė╔55.66%į÷┤¾ų┴82.96%.╩Y▄ŖĄ╚蹊┐▒Ē├„Ż¼«öĘ┤æ¬Ų„Ą─╚▌Ęežō║╔ė╔0.21 kg Īż(m3 Īżd)-1į÷┤¾ų┴0.84 kg Īż(m3 Īżd)-1ĢrŻ¼TNĄ─╚ź│²┬╩ė╔67.98%£pąĪų┴50.4%Ż¼┼c▒ŠčąŠ┐ĮY╣¹ėą▌^┤¾▓Ņ«ÉŻ¼┐╔─▄┼cīŹ“×čbų├║═Ę┤æ¬Śl╝■ėąĻP.īŹ“ץ─Ū░40ČÓ╠ņŻ¼│÷╦«ųąĄ─NH4+-N║═NO2--NØŌČ╚▌^ĘĆČ©Ż¼Ęųäeį┌0~30 mg ĪżL-1║═4~53 mg ĪżL-1ų«ķg▓©äė.«ö▀M╦«NH4+-N║═NO2--NØŌČ╚Ęųäeį÷┤¾ų┴240 mg ĪżL-1║═300 mg ĪżL-1ĢrŻ¼│÷╦«ųąĄ─NH4+-N║═NO2--NØŌČ╚ėą╦∙į÷┤¾Ż¼Ą½į÷┤¾Ę∙Č╚▌^ąĪŻ¼═©▀^ĮĄĄ═▀M╦«Ą─pHųĄŻ¼│÷╦«ųąĄ─NH4+-N║═NO2--NØŌČ╚Įø▀^1 dĘųäeĮĄĄ═ų┴0~10 mg ĪżL-1║═15~30 mg ĪżL-1ų«ķg.ļSų°▀M╦«NH4+-N║═NO2--NØŌČ╚Ą─└^└mį÷┤¾Ż¼«ö▀M╦«NH4+-N║═NO2--NØŌČ╚Ęųäe▀_ĄĮ330 mg ĪżL-1║═430mg ĪżL-1ĢrŻ¼Ę┤æ¬Ų„│÷╦«╦«┘|═╗╚╗É║╗»Ż¼│÷╦«Ą─NH4+-N║═NO2--NØŌČ╚Ęųäeį÷┤¾ų┴89 mg ĪżL-1║═163 mg ĪżL-1.┤╦ĢrNH4+-N║═NO2--NĄ─╚ź│²┬╩Ęųäe×ķ73.0%║═62.1%Ż¼┤╦ĢrģÆč§░▒č§╗»Š·Ą─╗Ņąį╩▄ĄĮęųųŲ. Dapena-MoraĄ╚蹊┐░l¼F«öNO2--NØŌČ╚▀_ĄĮ350mg ĪżL-1ĢrŻ¼ģÆč§░▒č§╗»Š·Ą─╗ŅąįĢ■ĮĄĄ═50%.į┌▓╗═¼čąŠ┐ųąŻ¼NO2--Nī”ģÆč§░▒č§╗»Š·Ą─ęųųŲØŌČ╚╩Ū▓╗═¼Ż¼▀@┐╔─▄┼cŠ·ĘN╝░▓┘ū„Śl╝■ėąĻP.×ķ┴╦▌^┐ņĄž╗ųÅ═Ę┤æ¬Ų„Ą─├ōĄ¬ąį─▄Ż¼īó▀M╦«NH4+-N║═NO2--NØŌČ╚ĘųäeĮĄĄ═ų┴310 mg ĪżL-1║═400 mg ĪżL-1.Įø▀^7 dĄ─╗ųÅ═Ų┌Ż¼│÷╦«ųąĄ─NH4+-N║═NO2--NØŌČ╚ĘųäeĮĄĄ═ų┴16 mg ĪżL-1║═56 mg ĪżL-1Ż¼NH4+-N║═NO2--NĄ─╚ź│²┬╩Ęųäe╗ųÅ═ų┴95.0%║═86.6%.蹊┐▒Ē├„Ż¼NO2--NČŠąį’@ų°ÅŖė┌NH4+-NŻ¼į┌▀M╦«NO2--NØŌČ╚▌^Ė▀╗“▀M╦«NO2--N/NH4+-NŽÓī”▌^Ė▀Ą─Śl╝■Ž┬Ż¼Ę┤æ¬Ų„ąį─▄ęūę“╗∙┘|ęųųŲČ°╩¦ĘĆ.
ĪĪĪĪ
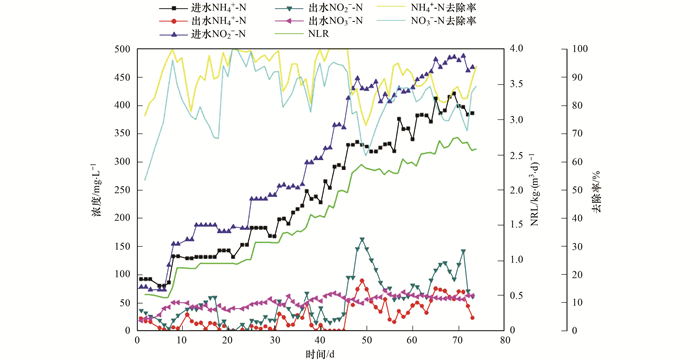
łD 2 ANAMMOXĘ┤æ¬Ų„Ą─├ōĄ¬ąį─▄
ĪĪĪĪ2.2 ANAMMOXĘ┤æ¬Ų„╬ó╔·╬’╚║┬õČÓśėąį蹊┐2.2.1 AlphaČÓśėąįųĖöĄĘų╬÷
ĪĪĪĪė╔Observed species║═Chao1ųĖöĄ┐╔ų¬ (ęŖ▒Ē 1)Ż¼Ė╗╝»ļAČ╬Ę┤æ¬Ų„ā╚Ą─╬ó╔·╬’╚║┬õŽÓī”žSČ╚ę¬Ė▀ė┌ĘĆČ©▀\ąąļAČ╬Ż¼Č°į┌ĘĆČ©▀\ąąļAČ╬Ż¼Ę┤æ¬Ų„ā╚Ą─╔Žīė╬ó╔·╬’╚║┬õŽÓī”žSČ╚ę¬Ė▀ė┌Ž┬īėŻ¼šf├„Ę┤æ¬Ų„ā╚Ą─╬ó╔·╬’Įø▀^ę╗Č╬ĢrķgĄ─±Z╗»║¾Ż¼▓┐Ęų╬ó╔·╬’▀mæ¬▓╗┴╦¤oÖCģÆ解hŠ│Č°ųØu╦└═÷Ż¼įņ│╔╬ó╔·╬’Ą─žSČ╚ĮĄĄ═.ė╔PD whole treeĪóShannon║═SimpsonųĖöĄ┐╔ų¬ (ęŖ▒Ē 1)Ż¼Ė╗╝»ļAČ╬Ę┤æ¬Ų„ā╚Ą─╬ó╔·╬’╚║┬õČÓśėąįę¬Ė▀ė┌ĘĆČ©▀\ąąļAČ╬Ż¼Č°ĘĆČ©▀\ąąļAČ╬Ę┤æ¬Ų„ā╚Ą─╔Žīė╬ó╔·╬’╚║┬õČÓśėąįę¬Ė▀ė┌Ž┬īėŻ¼šf├„Ę┤æ¬Ų„ā╚Ą─AnAOBĮø▀^ę╗Č╬ĢrķgĄ─±Z╗»║¾ą╬│╔┴╦ā×ä▌ĘN╚║Ż¼ī¦ų┬╬ó╔·╬’╚║┬õČÓśėąįĄ─£p╔┘.ė╔ė┌ÅU╦«╩ŪÅ─Ę┤æ¬Ų„Ą─Ąū▓┐▒├╚ļŻ¼ę“┤╦Ę┤æ¬Ų„Ž┬īėĄ─AnAOBūŅŽ╚▒╗±Z╗»│╔ā×ä▌ĘN╚║Ż¼ę“Č°Ž┬īėĄ─╬ó╔·╬’╚║┬õČÓśėąįꬥ═ė┌╔Žīė.
ĪĪ
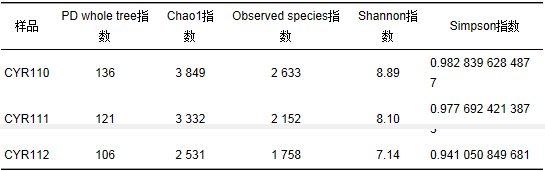
ĪĪĪĪ▒Ē 1 ╔·╬’ČÓśėąįĘų╬÷
ĪĪĪĪ2.2.2 ķT╦«ŲĮ╬’ĘNŽÓī”žSČ╚Ęų╬÷
ĪĪĪĪė╔łD 3┐╔ų¬Ż¼╦∙£yĄ├Ą─ŽÓī”žSČ╚┤¾ė┌Ą╚ė┌1%Ą─ķTļ`ī┘ė┌14éĆķT.ŲõųąŻ¼Ė╗╝»ļAČ╬Ę┤æ¬Ų„ųąĄ─╬ó╔·╬’ų„ę¬ęįūāą╬Š·ķTProteobacteriaĪóöMŚUŠ·ķTBacteroidetesĪóŠGÅØŠ·ķTChloroflexiĪó╚¬╣┼Š·ķTCrenarchaeota║═ÅV╣┼Š·ķTEuryarchaeota×ķų„Ż¼ĘĆČ©▀\ąąļAČ╬Ę┤æ¬Ų„ųąĄ─╬ó╔·╬’ų„ę¬ęįūāą╬Š·ķTProteobacteriaĪóöMŚUŠ·ķTBacteroidetesĪóŠGÅØŠ·ķTChloroflexi║═ĖĪ├╣Š·ķTPlanctomycetes×ķų„Ż¼Ę┤æ¬Ų„ųąĄ─ĖĪ├╣Š·ķT║═ūāą╬Š·ķTėą┴╦├„’@Ą─į÷╝ėŻ¼Ęųäeė╔1.1%Īó27.9%į÷╝ėĄĮ┴╦26.4%Īó39.9%.Č°öMŚUŠ·ķTĪóŠGÅØŠ·ķTĪó╚¬╣┼Š·ķT║═ÅV╣┼Š·ķTČ╝├„’@£p╔┘.Ę┤æ¬Ų„╔ŽŽ┬īė╬’ĘNŽÓī”žSČ╚ėą├„’@Ą─▓Ņ«É.Ąū▓┐Ą─ĖĪ├╣Š·ķT├„’@▒╚╔ŽīėČÓŻ¼Č°╔ŽīėĄ─öMŚUŠ·ķTę¬▒╚Ąū▓┐ČÓ.
ĪĪ
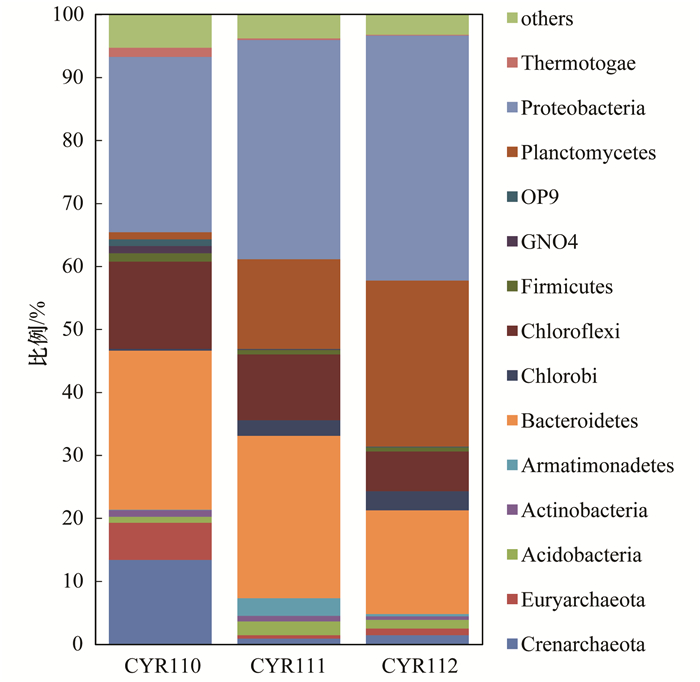
łD 3 ķT╦«ŲĮ╬’ĘNŽÓī”žSČ╚Ęų▓╝
ĪĪĪĪ2.2.3 ┐Ų╦«ŲĮ╬’ĘNŽÓī”žSČ╚Ęų╬÷
ĪĪĪĪė╔łD 4┐╔ų¬Ż¼╦∙£yĄ├Ą─ŽÓī”žSČ╚┤¾ė┌Ą╚ė┌1%Ą─┐Ųļ`ī┘ė┌21éĆ┐ŲŻ¼ŲõųąŻ¼Ė╗╝»ļAČ╬Ę┤æ¬Ų„ųąĄ─╬ó╔·╬’ų„ę¬ęįģÆ觎ēŠ·┐ŲAnaerolinaceaeĪó╝tŁhŠ·┐ŲRhodocyclaceaeĪóģ▓├½å╬µ▀Š·┐ŲComamonadaceae×ķų„.ĘĆČ©▀\ąąļAČ╬Ę┤æ¬Ų„ųąĄ─╔Žīė╬█─Óų„ę¬ęįģÆ觎ēŠ·┐ŲAnaerolinaceaeĪó╝tŁhŠ·┐ŲRhodocyclaceaeĪóģ▓├½å╬µ▀Š·┐ŲComamonadaceae║═Brocadiaceae×ķų„Ż¼Ž┬īė╬█─Óų„ę¬ęįģÆ觎ēŠ·┐ŲAnaerolinaceaeĪó╝tŁhŠ·┐ŲRhodocyclaceaeĪóģ▓├½å╬µ▀Š·┐ŲComamonadaceaeĪó³Så╬░¹Š·┐ŲXanthomonadaceae║═Brocadiaceae×ķų„.Ę┤æ¬Ų„ųąĄ─ģÆ觎ēŠ·┐ŲAnaerolinaceaeĪó╝ū═ķĮzĀŅŠ·┐ŲMethanosaetaceaeĪóWSA2ĪóĖ»┬▌ą²Š·┐ŲSaprospiraceaeĪó¤ßµ▀Š·┐ŲThermotogaceae║═╗źĀIŠ·┐ŲSyntrophaceaeėą├„’@Ą─£p╔┘Ż¼ŲõųąWSA2ĪóĖ»┬▌ą²Š·┐ŲSaprospiraceaeĪó¤ßµ▀Š·┐ŲThermotogaceae║═╗źĀIŠ·┐ŲSyntrophaceaeį┌ĘĆČ©▀\ąąļAČ╬Äū║§┐┤▓╗ĄĮ.Č°╝tŁhŠ·┐ŲRhodocyclaceaeĪóģ▓├½å╬µ▀Š·┐ŲComamonadaceaeĪó³Så╬░¹Š·┐ŲXanthomonadaceae║═Brocadiaceaeėą┴╦├„’@Ą─į÷╝ėŻ¼ŲõųąBrocadiaceaeį÷╝ėūŅ×ķ’@ų°Ż¼ė╔0.074 6%į÷╝ėĄĮ┴╦24.572 7%.Ę┤æ¬Ų„╔ŽīėBrocadiaceaeš╝12.486 9%Ż¼Ž┬īėš╝24.572 7%Ż¼Ž┬īėBrocadiaceaeĄ─žSČ╚├„’@▒╚╔ŽīėĖ▀. Brocadiaceaeī┘ė┌ĖĪ├╣Š·ķTŽ┬Ą─ę╗éĆ┐ŲŻ¼ęč│÷¼FĄ─ģÆč§░▒č§╗»Š·ų„ę¬üĒūįė┌▀@éĆ┐Ų.ė╔┤╦┐╔ų¬Ż¼ģÆč§░▒č§╗»Ę┤æ¬Ų„Įø▀^ę╗Č╬ĢrķgĄ─▀\ąą║¾Ż¼Ę┤æ¬Ų„ųąĄ─ģÆč§░▒č§╗»╬ó╔·╬’├„’@į÷╝ė┴╦.Č°Ę┤æ¬Ų„Ž┬īėĄ─ģÆč§░▒č§╗»╬ó╔·╬’├„’@▒╚╔ŽīėĖ▀.
ĪĪĪĪ
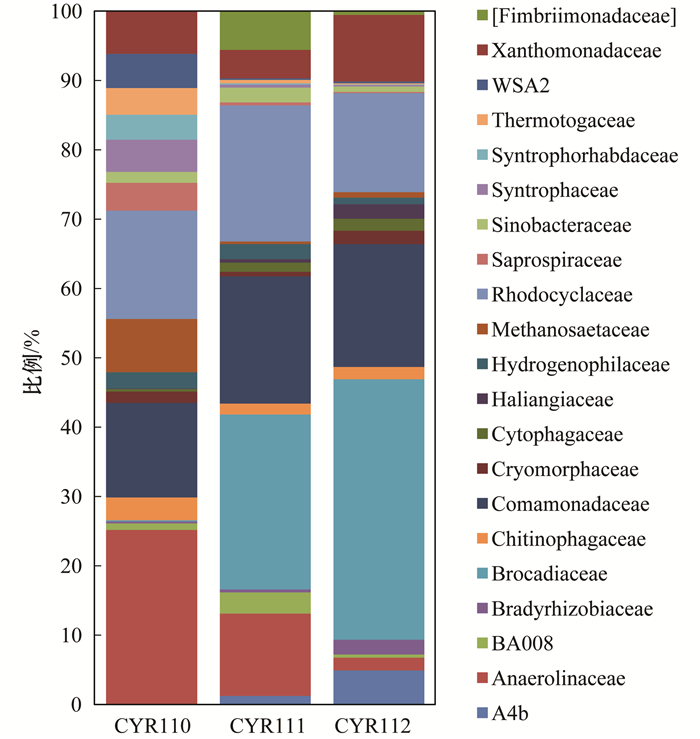
łD 4 ┐Ų╦«ŲĮ╬’ĘNŽÓī”žSČ╚Ęų▓╝
ĪĪĪĪ2.2.4 ╬’ĘNŽÓī”žSČ╚¤ßłDĘų╬÷
ĪĪĪĪė╔łD 5┐╔ų¬Ż¼ĘĆČ©▀\ąąļAČ╬Ą─Ž┬īė╬█─ÓųąĄ─BrocadiaceaeĪó╩╔└wŠSŠ·┐ŲCytophagaceaeĪóģ▓├½å╬░¹Š·┐ŲComamonadaceaeĪóSB1Īó³Så╬░¹Š·┐ŲXanthomonadaceaeĪó┬²╔·Ė∙┴÷Š·┐ŲBradyrhizobiaceaeĪóA4bĪóHaliangiaceaeĪóCryomorphaceaeĪó╔·Įz╬óŠ·┐ŲHyphomicrobiaceae╚║┬õŽÓī”žSČ╚ę¬▒╚╔ŽīėĖ▀Ż¼ŲõųąŻ¼╔Žīė╬█─ÓųąĄ─BrocadiaceaeĪó╩╔└wŠSŠ·┐ŲCytophagaceaeĪóģ▓├½å╬░¹Š·┐ŲComamonadaceaeĪóSB1╚║┬õŽÓī”žSČ╚ę¬▒╚Ė╗╝»ļAČ╬Ė▀.ŽÓĘ┤Ż¼Ė╗╝»ļAČ╬Ę┤æ¬Ų„ųąĄ─¤ß┼█Š·┐ŲThermotogaceaeĪó╗źĀIŚUŠ·┐ŲSyntrophobacteraceaeĪóSyntrophorhabdaceaeĪó╗źĀIŠ·┐ŲSyntrophaceaeĪóĖ»┬▌ą²Š·┐ŲSaprospiraceaeĪóWSA2Īó╝ū═ķĮzĀŅŠ·┐ŲMethanosaetaceaeĪó╝ū═ķŚUŠ·┐ŲMethanobacteriaceaeĪóģÆ觎ēŠ·┐ŲAnaerolinaceaeĪó┼»ŽēŠ·┐ŲCaldilineaceaeĪó╚ŌŚUŠ·┐ŲCarnobacteriaceae╚║┬õŽÓī”žSČ╚ę¬▒╚ĘĆČ©▀\ąąļAČ╬Ė▀.┐é¾wüĒ┐┤Ż¼Ė╗╝»ļAČ╬Ę┤æ¬Ų„ā╚Ą─╬ó╔·╬’╚║┬õĮYśŗ┼cĘĆČ©▀\ąąļAČ╬ŽÓ▒╚ŽÓ▓Ņ║▄┤¾Ż¼Č°ĘĆČ©▀\ąąļAČ╬Ę┤æ¬Ų„╔ŽŽ┬īėĄ─╬ó╔·╬’╚║┬õĮYśŗę▓ėą▌^┤¾Ą─▓Ņ«É. ChenĄ╚蹊┐▒Ē├„ABRģÆč§░▒č§╗»Ę┤æ¬Ų„Ė„Ė¶╩ęĄ─╬ó╔·╬’╚║┬õĮYśŗ┤µį┌▌^┤¾Ą─▓Ņ«É, ┼c▒ŠčąŠ┐ĮY╣¹ŽÓĘ¹.
ĪĪ
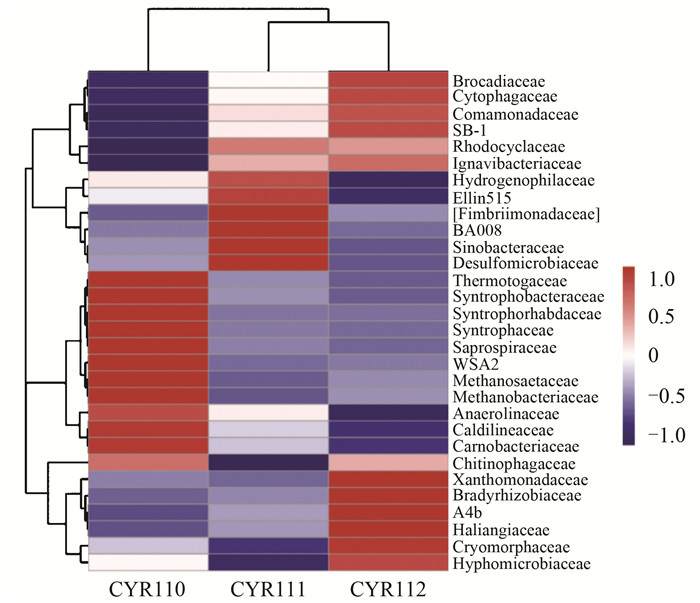
łD 5 ╬’ĘNŽÓī”žSČ╚Š█ŅÉ (┐Ų╦«ŲĮ)
ĪĪĪĪ2.2.5 ├ōĄ¬╝ÜŠ·ČÓśėąįĘų╬÷
ĪĪĪĪ═©▀^Ė▀═©┴┐Ą─öUį÷ūė£yą“Ż¼╣▓ĶbČ©│÷ķTĄ─╝ÜŠ·40ĘN.ŲõųąŻ¼╔µ╝░ĄĮ├ōĄ¬╝ÜŠ·Ą─ėąūāą╬Š·ķTĪóĖĪ├╣Š·ķT║═Ž§╗»┬▌ą²Š·ķT3ĘNŻ¼ūāą╬Š·ķTžSČ╚ūŅĖ▀Ż¼┼cČÓöĄģÆč§░▒č§╗»Ę┤æ¬Ų„╬ó╔·╬’╚║┬õĘų▓╝ŪķørŽÓę╗ų┬.
ĪĪĪĪį┌ūāą╬Š·ķTųąŻ¼Ė╗╝»ļAČ╬Ą─śėŲĘųą╣▓ĶbČ©│÷ī┘Ą─╝ÜŠ·67ĘNŻ¼š╝ō■▒╚└²▌^Ė▀Ą─╩Ū╠šČ“╩ŽŠ·ī┘Thauera(1.532%)ĪóDok59(1.318%)ĪóThibacillus┴“ŚUŠ·ī┘ (0.888%) ║═╗źĀIŚUŠ·ī┘Syntrophobacter(0.650%).ĘĆČ©▀\ąąļAČ╬Ą─╔Žīė╬█─ÓśėŲĘųą╣▓ĶbČ©│÷ī┘Ą─╝ÜŠ·56ĘNŻ¼š╝ō■▒╚└²▌^Ė▀Ą─╩ŪDok59ī┘ (4.644%)ĪóThaueraī┘ (2.560%)ĪóThibacillusī┘ (1.074%) ║═├ō┴“╬óŠ·ī┘Desulfomicrobium(0.923%)Ż¼ĘĆČ©▀\ąąļAČ╬Ą─Ž┬īė╬█─ÓśėŲĘųą╣▓ĶbČ©│÷ī┘Ą─╝ÜŠ·54ĘNŻ¼š╝ō■▒╚└²▌^Ė▀Ą─╩ŪDok59ī┘ (6.014%)ĪóThaueraī┘ (1.447%)ĪóThibacillusī┘ (0.638%) ║═Thermomonasī┘ (0.503%).ūāą╬Š·ķTųąī┘Ą─╝ÜŠ·ČÓśė£pąĪ. Thaueraī┘ĪóDok59ī┘║═Thibacillusī┘žSČ╚ūŅĖ▀Ż¼ŲõųąThaueraī┘║═Thibacillusī┘╩Ū”┬-ūāą╬Š·ŠVųąĄ─Ę┤Ž§╗»╝ÜŠ·.┤╦═ŌŻ¼Ė╗╝»ļAČ╬Ą─śėŲĘųąų╗Öz£y│÷Nitrosovibrio▀@ę╗ĘN░▒č§╗»╝ÜŠ· (AOB)Ż¼āHš╝0.011%Ż¼ī┘ė┌”┬-ūāą╬Š·üåŠV. AOBī┘ė┌─▄ē“į┌ėąč§Śl╝■Ž┬īó░▒č§╗»×ķü厧╦ß¹}Ą─╗»─▄¤oÖCūįBą═╝ÜŠ·.▒ŠīŹ“×čbų├ø]ėąīŻķTĄ─│²č§įOéõŻ¼ę“┤╦Ģ■░ķļSėą╔┘┴┐Ą─č§ÜŌ▀M╚ļĘ┤æ¬Ų„ųą×ķAOB╠ß╣®ėąč§ŁhŠ│.ė╔łD 6┐╔ęį┐┤│÷Ż¼”żNO2--N/”żNH4+-NųĄį┌ūŅ│§Ą─10 dė╔0.60į÷┤¾ų┴1.31Ż¼╚╗║¾į┌1.05~1.41ų«ķg▓©äė. ”żNO3--N/”żNH4+-NųĄį┌ūŅ│§Ą─10 dė╔0.22į÷┤¾ų┴0.44Ż¼╚╗║¾ųØu£pąĪų┴0.15.šf├„Ė╗╝»ļAČ╬▓┐ĘųNH4+-N▒╗AOB▐D╗»×ķü厧╦ß¹}.Č°ĘĆČ©▀\ąąļAČ╬Ą─”żNO2--N/”żNH4+-NųĄ▌^ĮėĮ³└ĒšōųĄ1.32Ż¼šf├„┤╦ĢrNH4+-NÄū║§╚½▓┐ė╔AnAOB╦∙└¹ė├.
ĪĪ

łD 6 3ĘNĄ¬╦žĄ─▒╚ųĄūā╗»
ĪĪĪĪŽ§╗»┬▌ą²Š·ķT╩Ū£yą“Ą├ĄĮĄ─╚²┤¾├ōĄ¬╝ÜŠ·ķTųą╦∙š╝▒╚└²ūŅąĪĄ─Ż¼į┌0.035%~0.188%ų«ķgŻ¼Ė╗╝»ļAČ╬Ą─║¼┴┐ūŅĖ▀ (0.188%).═Ų£yĖ╗╝»ļAČ╬Ą─AOBīó▓┐Ęų░▒▐D╗»×ķü厧╦ß¹}Ż¼ī¦ų┬╩ŻėÓü厧╦ß¹}Ą─Ęe└█Ż¼Č°Ž§╗»┬▌ą²Š·ķT╩Ūü厧╗»Ę┤æ¬Ą─ų„ę¬╬ó╔·╬’Ż¼ę“┤╦║¼┴┐Ė▀ė┌ĘĆČ©▀\ąąļAČ╬.į┌Ž§╗»┬▌ą²Š·ķTųąŻ¼╣▓ĶbČ©│÷2ĘNī┘Ą─╝ÜŠ·Ż¼Ęųäe×ķGOUTA19║═NitrospiraŻ¼ŲõųąŻ¼Nitrospira×ķü厧¹}č§╗»╝ÜŠ· (NOB)Ż¼╦∙š╝▒╚└²į┌0.023%~0.174%ų«ķgŻ¼Ė╗╝»ļAČ╬Ą─║¼┴┐ūŅĖ▀ (0.174%).═Ų£yĖ╗╝»ļAČ╬Ą─AOBīó▓┐Ęų░▒▐D╗»×ķü厧╦ß¹}Ż¼ī¦ų┬╩ŻėÓü厧╦ß¹}Ą─Ęe└█Ż¼Č°NOB┐╔īóü厧╦ß¹}č§╗»×ķŽ§╦ß¹}Ż¼ę“┤╦║¼┴┐Ė▀ė┌ĘĆČ©▀\ąąļAČ╬.
ĪĪĪĪį┌ĖĪ├╣Š·ķTųą╣▓ĶbČ©│÷ī┘Ą─╝ÜŠ·5ĘNŻ¼Ęųäe×ķGemmatiaĪóA17ĪóPirellulaĪóPlanctomycesĪóCandidatus brocadiaŻ¼ŲõųąŻ¼Candidatus brocadia╦∙š╝Ą─▒╚└²ūŅĖ▀Ż¼į┌0.011%~1%ų«ķg.ĘĆČ©▀\ąąļAČ╬Candidatus brocadia╦∙š╝Ą─▒╚└² (1%) Ė▀ė┌Ė╗╝»ļAČ╬ (0.011%)Ż¼┼cĘĆČ©▀\ąąļAČ╬Ą─╚ź│²žō║╔1.04kg Īż(m3 Īżd)-1Ė▀ė┌Ė╗╝»ļAČ╬0.29 kg Īż(m3 Īżd)-1ŽÓę╗ų┬. Candidatus brocadiaī┘╩ŪĄ┌ę╗éĆ▒╗Ė╗╝»ĶbČ©Ą─AnAOB[25]Ż¼ī┘ė┌Brocadiaceae┐Ų.Č°ī┘ė┌Brocadiaceae┐ŲĄ─96%ą“┴ą═©▀^BLAST▀Mąąą“┴ą▒╚ī”░l¼F┼cČÓŚlą“┴ą═¼į┤ąįŽÓ╦ŲŻ¼ę“┤╦¤oĘ©┤_Č©╩Ū──ĘNī┘.═Ų£y╬┤ĘųŅÉŠ·ī┘ųą┐╔─▄║¼ėą╬┤ĶbČ©│÷Ą─AnAOBą┬ĘN.Š▀¾wģóęŖ╬█╦«īÜ╔╠│Ū┘Y┴Ž╗“http://www.jianfeilema.cnĖ³ČÓŽÓĻP╝╝ąg╬─ÖnĪŻ
ĪĪĪĪ3 ĮYšō
ĪĪĪĪ(1) ▓╔ė├╠ßĖ▀▀M╦«NH4+-N║═NO2--NØŌČ╚Ą─ĘĮ╩ĮīóANAMMOXĘ┤æ¬Ų„Ą─╚▌Ęežō║╔ė╔0.52 kg Īż(m3 Īżd)-1į÷┤¾ų┴2.75 kg Īż(m3 Īżd)-1Ż¼NH4+-NĪóNO2--N║═TNĄ─╚ź│²┬╩ĘųäeÅ─76.18%Īó53.47%Īó55.66%į÷┤¾ų┴94.04%Īó86.97%Īó82.96%Ż¼AnAOB│╔╣”Ė╗╝».
ĪĪĪĪ(2) ė╔AlphaČÓśėąįųĖöĄĘų╬÷┐╔ų¬Ż¼Ę┤æ¬Ų„Ė╗╝»ļAČ╬Ą─╬ó╔·╬’╚║┬õŽÓī”žSČ╚║═╚║┬õČÓśėąįę¬Ė▀ė┌ĘĆČ©▀\ąąļAČ╬Ż¼Č°į┌ĘĆČ©▀\ąąļAČ╬Ż¼Ę┤æ¬Ų„ā╚Ą─╔Žīė╬ó╔·╬’╚║┬õŽÓī”žSČ╚║═╚║┬õČÓśėąįę¬Ė▀ė┌Ž┬īė.
ĪĪĪĪ(3) ė╔ķT╦«ŲĮ╬’ĘNŽÓī”žSČ╚Ęų╬÷┐╔ų¬Ż¼Ę┤æ¬Ų„ųąĄ─ĖĪ├╣Š·ķT║═ūāą╬Š·ķTėą├„’@Ą─į÷╝ėŻ¼ŲõųąŻ¼ĖĪ├╣Š·ķTĄ─žSČ╚į÷┤¾ūŅ×ķ’@ų°.Ę┤æ¬Ų„Ąū▓┐Ą─ĖĪ├╣Š·ķT├„’@▒╚╔ŽīėČÓŻ¼Č°╔ŽīėĄ─öMŚUŠ·ķTę¬▒╚Ąū▓┐ČÓ.ė╔┐Ų╦«ŲĮ╬’ĘNŽÓī”žSČ╚Ęų╬÷┐╔ų¬Ż¼Ę┤æ¬Ų„Ė╗╝»ļAČ╬ęįģÆ觎ēŠ·┐ŲĄ─ŽÓī”žSČ╚ūŅĖ▀Ż¼×ķ9.47%.ĘĆČ©▀\ąąļAČ╬BrocadiaceaeĄ─ŽÓī”žSČ╚ūŅĖ▀Ż¼▀_ĄĮ┴╦24.57%Ż¼│╔×ķā×ä▌Š·╚║.
ĪĪĪĪ(4) ė╔╬’ĘNŽÓī”žSČ╚¤ßłDĘų╬÷┐╔ų¬Ż¼Ė╗╝»ļAČ╬Ą─╬ó╔·╬’╚║┬õĮYśŗ┼cĘĆČ©▀\ąąļAČ╬ŽÓ▒╚ŽÓ▓Ņ║▄┤¾Ż¼Č°ĘĆČ©▀\ąąļAČ╬Ę┤æ¬Ų„╔ŽŽ┬īė╬ó╔·╬’╚║┬õĮYśŗę▓ėą▌^┤¾Ą─▓Ņ«É.
ĪĪĪĪ(5) ė╔£yą“ĮY╣¹┐╔ų¬Ż¼ĘĆČ©▀\ąąļAČ╬UBFģÆč§░▒č§╗»Ę┤æ¬Ų„ųąĄ─├ōĄ¬╝ÜŠ·▌^×ķžSĖ╗Ż¼Ųõųąūāą╬Š·ķTĪóĖĪ├╣Š·ķT║═Ž§╗»┬▌ą²Š·ķTĘųäeš╝39.9%Īó26.4%║═0.188%.ūāą╬Š·ķTųąš╝ō■▒╚└²▌^Ė▀Ą─╩ŪDok59ī┘ĪóThaueraī┘ĪóThibacillusī┘║═Thermomonasī┘.ĖĪ├╣Š·ķTų„ę¬░³║¼Candidatus brocadia.

