ļp(3-░▒╗∙▒¹╗∙)░Ęī”╗ņ║ŽŠ·┬õ╔·╬’─żą╬│╔ęųųŲ╝░ĮŌ¾wą¦æ¬ÖCųŲ蹊┐
ųąć°╬█╦«╠Ä└Ē╣ż│╠ŠW ĢrķgŻ║2018-7-12 10:12:21
╬█╦«╠Ä└Ē╝╝ąg | ģRŠ█╚½Ū“Łh▒Ż┴”┴┐Ż¼ĮĄĄ═Ų¾śIų╬╬█│╔▒Š
ĪĪĪĪ1 ę²čį(Introduction)
ĪĪĪĪ╔·╬’─ż(Biofilm)╩Ūė╔ĖĮų°ė┌ČĶąį╗“š▀╗Ņąį╣╠¾w▓─┴Ž▒Ē├µĄ─╬ó╔·╬’║═ė╔╬ó╔·╬’ūį╔ĒĘų├┌Ą─░¹═ŌŠ█║Ž╬’(Extracellular polymeric substances, EPS)╦∙ą╬│╔Ą─Ė▀Č╚ĮYśŗ╗»╬ó╔·╬’╚║┬õ(Costerton et al., 1999).╦³Ą─┤µį┌┐╔ę²Ųę╗ŽĄ┴ąĄ─ŁhŠ│å¢Ņ}, ╚ń─ż╬█╚ŠĪó╣▄Ą└Č┬╚¹ĪóĮī┘▒Ē├µĖ»╬g║═Ž¹ČŠą¦┬╩ĮĄĄ═Ą╚(Gino et al., 2010; Schwering et al., 2013).─┐Ū░ī”Ųõ┐žųŲČÓ▓╔ė├ÜóŠ·ä®Īó┐╣Š·ä®╗“Ųõ╦³┤¾ĘųūėęųŠ·ä®(Rosmaninho et al., 2007; Simões et al., 2010).Ą½▀@ą®╬’┘|Ą─ßīĘ┼¤oę╔ĦüĒ┴╦ą┬Ą─ŁhŠ│’LļU, ķLŲ┌╩╣ė├┐╣╔·╦ž╗“š▀ÜóŠ·ä®▀ĆĢ■ī¦ų┬╝ÜŠ·Ą─┐╣╦Äąįį÷╝ė, ─═╦Ä╝ÜŠ·┬¹čė(Davies, 2003).
ĪĪĪĪ└¹ė├╗»īWą┼╠¢ąĪĘųūė╬’┘|═©▀^ęųųŲ╬ó╔·╬’╗Ņąį, š{╣Ø╔·╬’─żĄ─░lš╣▀^│╠╩Ū─┐Ū░╔·╬’─żŅIė“Ą─蹊┐¤ß³c.ļp(3-░▒╗∙▒¹╗∙)░Ę╩Ū─│ą®╬ó╔·╬’ūį╔Ē«a╔·Ą─┐╔ęįę²Ųę╗ą®╔·╬’─żĮŌ¾wĄ─ą┼╠¢Ęųūė(Qu et al., 2016), ╠Įėæ▀@ĘNąĪĘųūė╬’┘|Ą─╔·╬’─żš{┐žū„ė├ī”ė┌ķ_░lą┬┼d╔·╬’─ż┐žųŲ╝╝ągŠ▀ėąųžę¬ęŌ┴x.─┐Ū░, ļp(3-░▒╗∙▒¹╗∙)░Ęī”╬ó╔·╬’ĖĮų°Ą─ęųųŲ蹊┐▌^╔┘ŪęČÓ╝»ųąė┌╝āŠ·╔·╬’─ż, ī”╗ņ║ŽŠ·┬õ╔·╬’─żĄ─ęųųŲ蹊┐§rėął¾Ą└.īŹļHŁhŠ│ųąĄ─╔·╬’─żČÓė╔╗ņ║ŽŠ·╚║ĮM│╔, ę“┤╦ėą▒žę¬čąŠ┐ļp(3-░▒╗∙▒¹╗∙)░Ęī”╗ņ║ŽŠ·┬õ╔·╬’─żęųųŲ╝░ĮŌŠ█ą¦æ¬.▒ŠčąŠ┐īóęįļp(3-░▒╗∙▒¹╗∙)░Ę×ķ─┐ś╦╬’┘|, ┐╝▓ņ▀@ĘNąĪĘųūė╬’┘|ī”╗ņ║ŽŠ·┬õ╔·╬’─żą╬│╔ęųųŲ╝░ĮŌ¾wą¦æ¬ÖCųŲ, ▓ó▀Mę╗▓Įī”Ųõ£pŠÅ─ż▒Ē├µ╔·╬’╬█╚ŠĄ─┐╔ąąąį▀Mąą╠Įėæ.įō蹊┐ī”ė┌Į©┴ó╗∙ė┌ąĪĘųūė╬’┘|Ą─╔·╬’─żęųųŲ╝░─ż▒Ē├µ╔·╬’╬█╚Š┐žųŲĘĮĘ©Š▀ėą└ĒšōārųĄ┼cīŹļHųĖī¦ęŌ┴x.
ĪĪĪĪ2 ▓─┴Ž┼cĘĮĘ©(Materials and methods)2.1 ╗»īWįćä®║═╬ó╔·╬’
ĪĪĪĪļp(3-░▒╗∙▒¹╗∙)░Ę┘Åūį├└ć°Sigma-Aldrich╣½╦Š, ╝āČ╚×ķ99%.Ųõ╦¹╗»īWįćä®┘Åė┌ē¶Ō∙├└╔·╬’┐Ų╝╝ėąŽ▐╣½╦Š(▒▒Š®, ųąć°).
ĪĪĪĪ╗ņ║ŽŠ·┬õüĒį┤ė┌ŪÕ║ė╬█╦«╠Ä└ĒÅS(▒▒Š®, ųąć°)Ą─╗Ņąį╬█─Ó, Įø▀^╔Ž║Ż├└╝¬╔·╬’ßt╦Ä┐Ų╝╝ėąŽ▐╣½╦Š═©▀^MiSeqŽĄĮyĘų╬÷ĮM│╔ų„ę¬░³└©ģ▓├½å╬░¹Š·ī┘(Comamonas)Īóµ▀¶~Š·┐Ų(Nakamurella)Īó─cŚUŠ·┐Ų(Enterobacteriaceae)Īó╦¾ĀŅč┐µ▀ŚUŠ·(Clostridium)ĪóŪ╩ų¼å╬░¹Š·ī┘(Sphingomonas)Īó╝┘å╬░¹Š·ī┘(Pseudomonas)Īó╣╠Ą¬┬▌Š·ī┘(Azospira)Īó╣čBå╬░¹Š·(Stenotrophomonas)║═ĶFŚUŠ·ī┘(Ferribacterium).╗Ņąį╬█─Ó╩ūŽ╚ė├┴ū╦ß¹}ŠÅø_╚▄ę║(PBS)ø_Ž┤3▒ķęį╚ź│²╬█─Ó╚▄ę║ųąĄ─EPS╝░ėąÖC╬’, ╚╗║¾īó╗Ņąį╬█─Óųžæęė┌║Ž│╔ÅU╦«ųąė├ė┌ųŲéõ╗ņ║ŽŠ·Š·æęę║(100 mg VSSĪżL-1)(┤¾╝s108 CFUĪżmL-1).║Ž│╔ÅU╦«░³║¼(mgĪżL-1)Ż║COD (ęįŲŽ╠č╠Ūėŗ╦Ń) 825;NH4Cl 192;KH2PO4 35.1;NaCl 100;MgSO4 100;CaCl2 10;pH=6.5~7.5.
ĪĪĪĪ2.2 ļp(3-░▒╗∙▒¹╗∙)░Ęī”╗ņ║ŽŠ·╚║╔·╬’─żą╬│╔ė░Ēæ蹊┐
ĪĪĪĪīó║¼ėą▓╗═¼ØŌČ╚ļp(3-░▒╗∙▒¹╗∙)░ĘĄ─Š·æęę║╝ė╚ļĄĮŠ█▒ĮęꎮĄ─12┐ū░Õųą, ├┐┐ū4 mL, ļp(3-░▒╗∙▒¹╗∙)░ĘĮKØŌČ╚Ęųäe×ķ0Īó2.62Īó6.55Īó13.10Īó26.20Īó65.50╗“131 mgĪżL-1, ├┐éĆØŌČ╚įOų├6éĆŲĮąąśėęį┤_▒ŻīŹ“ץ─£╩┤_ąį.īó╝ė═ĻśėŲĘĄ─12┐ū░Õų├ė┌30 Īµ║Ń£ž┼ÓBŽõųąĘųäeņoų╣┼ÓB12 h║═24 h.┼ÓB═Ļ│╔║¾ĖĮų°Ą─╔·╬’─ż┴┐└¹ė├ĮYŠ¦ūŽ╚Š╔½Ę©ŽÓī”Č©┴┐(Xiong et al., 2013).▒ŠčąŠ┐ęį╗Ņąį╬█─Óū„×ķ╔·╬’─żą╬│╔Ą─╗ņ║ŽŠ·üĒį┤, ļSų°╔·╬’─żĄ─ą╬│╔║═░lš╣, Ųõ╚║┬õĮYśŗ┐╔─▄Ģ■┼c╗Ņąį╬█─Óėąę╗ą®▓Ņäe, Ą½▒ŠčąŠ┐ųž³c▓╗╩ŪĻPūóŲõ╚║┬õĮYśŗč▌ūā▀^│╠, Č°╩Ūęį╔·╬’─żą╬╩Įžō▌dĄĮ╗∙Ąū▓─┴Ž▒Ē├µ╔·╬’┴┐Ą─ūā╗», ▓óęįŲõüĒįuār╔·╬’─żą╬│╔ęųųŲą¦æ¬.
ĪĪĪĪ2.3 ▓╗═¼╗∙Ąūīė╔Žļp(3-░▒╗∙▒¹╗∙)░Ęī”╗ņ║ŽŠ·╚║ĖĮų°ė░Ēæ
ĪĪĪĪęųųŲä®ī”╬ó╔·╬’ĖĮų°╠žąįĄ─ė░Ēæ║═ĖĮų°╗∙Ąū▓─┴Ž▒Ē├µ╠žąį├▄ŪąŽÓĻP, ╦∙ęį▒ŠčąŠ┐▀Mę╗▓Į┐╝▓ņ┴╦ļp(3-░▒╗∙▒¹╗∙)░Ęī”╬ó╔·╬’į┌▓╗═¼▓─┘|╗∙Ąūīė▒Ē├µĄ─ĖĮų°ė░Ēæ.Ęųäe▀xō±Š▀ėą▓╗═¼ėH╩Ķ╦«ąį╠žąįĄ─Š█▒Įęꎮ(6┐ū░Õ)Īó▓Ż┴¦Ų¼║═Š█╝ū╗∙▒¹Ž®╦ß╝ū§ź3ĘN▓─┴Ž×ķĖĮų°Įń├µ, 3ĘN▒Ē├µĮėė|ĮŪĘųäe×ķ93.4ĪŃĪó22.0ĪŃ║═81.9ĪŃ.Ęųäeīó20 mmĪ┴ 20 mm▓Ż┴¦Ų¼╗“Š█╝ū╗∙▒¹Ž®╦ß╝ū§źŲ¼ų├ė┌6┐ū░Õųą▀MąąĖĮų°īŹ“×, ├┐éĆ┐ū░Õųą╝ė╚ļ6 mL╬█─Óæęę║, į┘Ęųäe╝ė╚ļļp(3-░▒╗∙▒¹╗∙)░ĘĮKØŌČ╚×ķ0╗“131 mgĪżL-1.╬█─Óæęę║Īó╗∙┘|ØŌČ╚╝░▓┘ū„Śl╝■║═╔Ž╩÷Ą─ĖĮų°īŹ“×ę╗ų┬.┼ÓB24 h║¾, īó▌d▓ŻŲ¼║µĖ╔, ╚Š╔½▓óė├OlympusŽÓÖC▀Mąą┼─ššėøõø.
ĪĪĪĪ2.4 ļp(3-░▒╗∙▒¹╗∙)░Ęī”─ż╬█╚ŠŠÅĮŌą¦æ¬čąŠ┐
ĪĪĪĪīóļp(3-░▒╗∙▒¹╗∙)░Ę═Č╝ėĄĮ╚▄ę║═©▀^║═æęĖĪ╬ó╔·╬’ū„ė├ų«║¾, ╬ó╔·╬’į┌─ż▒Ē├µĄ─ĖĮų°īŹ“×ĘųäeęįņoæBĖĮų°║═äėæB╦└Č╦▀^×Vą╬╩Į┐╝▓ņ.ņoæBīŹ“×╚ńŽ┬Ż║īóų▒ÅĮ×ķ3.5 cmĄ──żŲ¼ų├ė┌6┐ū┼ÓB░Õ, ╝ė╚ļ10 mL╗ņ║ŽŠ·ę║, ═¼Ģr╝ė╚ļļp(3-░▒╗∙▒¹╗∙)░Ę╩╣ŲõĮKØŌČ╚Ęųäe×ķ0╗“131 mgĪżL-1, 30 Īµ║Ń£ž┼ÓBŽõųąņoų╣┼ÓB24 h, ╚Ī│÷─żŲ¼Ę┼į┌15 mL PBSųą│¼┬Ģ╩╣ĖĮų°Ą─╬ó╔·╬’═Ļ╚½├ō┬õ║¾į┌ą²£u╗ņ║ŽŲ„ųą┐ņ╦┘╗ņ║Ž15 s, £yČ©╝ÜŠ·OD600ųĄ(Åł└¹ŲĮĄ╚, 2011)(ę“ĮYŠ¦ūŽ╚Š┴ŽĢ■║═─ż▓─┴Ž▒Ē├µĮY║Žė░ĒæīŹ“×ĮY╣¹, ╣╩▓╔ė├╗ņ║Ž£uą²├ōļx╔·╬’─ż║¾ų▒Įė£yŠ·¾w╣Ō├▄Č╚Ą─ĘĮĘ©).äėæB▀^×V▓╔ė├║Ń┴„▀^×V─Ż╩Į, ╩╣ė├ę╗┼_╚õäė▒├īóā”╦«╚▌Ų„ųą╗ņ║ŽŠ·ę║(║¼ėą╗“▓╗║¼ėą131 mgĪżL-1Ą─ļp(3-░▒╗∙▒¹╗∙)░Ę)═©▀^╣▄Ą└ē║╚ļ─żĮM╝■(─żų▒ÅĮ4 cm), ļŖūė╠ņŲĮ╩š╝»╚▄ę║Ą─┘|┴┐öĄō■, «ö─ż▒Ē├µįņ│╔ę╗Č©Č┬╚¹ų«║¾, ─żē║į÷╝ė, ē║┴”▒ĒūxöĄ╔Ž╔².═©▀^ē║┴”▒ĒūxöĄ┐╔ķgĮėįuār─ż╬█╚Š░lš╣ĀŅør.
ĪĪĪĪ2.5 ļp(3-░▒╗∙▒¹╗∙)░Ęī”╗ņ║ŽŠ·╚║╔·╬’─żĮŌ¾wė░Ēæ蹊┐
ĪĪĪĪ×ķ┴╦┐╝▓ņļp(3-░▒╗∙▒¹╗∙)░Ęī”╔·╬’─żĄ─ĮŌ¾wąį─▄, ąĶŅAŽ╚┼ÓB╔·╬’─ż(12 h).╚╗║¾īó▌d▓ŻŲ¼─├│÷Ž┤Ą¶╬┤ĖĮų°Ą─╬ó╔·╬’.╚╗║¾Ę┼ų├ė┌║¼ėą131 mgĪżL-1ļp(3-░▒╗∙▒¹╗∙)░ĘĄ─PBS╚▄ę║ųą, ┴Ē═ŌĄ─▌d▓ŻŲ¼Ę┼╚ļų╗║¼ėąPBSĄ─╚▄ę║ųąū„×ķī”šš, 30 ĪµĘųäeņoų├6Īó8Īó10╗“12 h║¾, īó▌d▓ŻŲ¼─├│÷Ž┤Ą¶æęĖĪĄ─╬ó╔·╬’, ╩ŻėÓ╔·╬’┴┐▓╔ė├╚ń╔ŽĮYŠ¦ūŽ╚Š╔½Ę©▀MąąŽÓī”Č©┴┐.
ĪĪĪĪ2.6 ļp(3-░▒╗∙▒¹╗∙)░Ęī”æęĖĪ╬ó╔·╬’EPSĘų▓╝ė░Ēæ蹊┐
ĪĪĪĪ×ķ┴╦╠Įėæļp(3-░▒╗∙▒¹╗∙)░Ęī”╔·╬’─żą╬│╔ęųųŲÖC└Ē, ▒ŠčąŠ┐▀Mę╗▓Įī”æęĖĪ╗ņ║ŽŠ·╚║╬ó╔·╬’Ą─EPS▀Mąą┐╝▓ņ.Š▀¾w▓┘ū„▓Į¾E╚ńŽ┬Ż║īó║¼ėąę╗Č©ØŌČ╚Ą─╬█─Óæęę║║═╗∙┘|(╚ń╔Ž)ų├ė┌50 mLļxą─╣▄ųą, ═¼Ģr╝ė╚ļļp(3-░▒╗∙▒¹╗∙)░Ę╩╣ŲõūŅĮKØŌČ╚Ęųäe×ķ0Īó131 mgĪżmL-1, 30 ĪµĘ┼ų├ę╗Č©Ģrķg, £yČ©EPS║¼┴┐ūā╗», ═¼Ģr£yČ©æęĖĪę║Ą─OD600ūā╗».EPSĄ─╠ß╚Īģó┐╝XuanĄ╚(2010)Ą─ĘĮĘ©, Ą░░ū£yČ©▓╔ė├Ą─LowryĘ©(Frølund et al., 1996), ČÓ╠ŪĄ─£yČ©▓╔ė├Ą─╩Ū┴“╦ß-▒ĮĘė▒╚╔½Ę©(Dubois et al., 1956);eDNAĄ─£yČ©ģó┐╝Allesen-HolmĄ╚(2006)Ą─ĘĮĘ©.
ĪĪĪĪ3 ĮY╣¹┼cėæšō(Results and discussion)3.1 ļp(3-░▒╗∙▒¹╗∙)ī”╗ņ║ŽŠ·╚║╔·╬’─żą╬│╔ė░Ēæ
ĪĪĪĪ▓╗═¼ØŌČ╚ļp(3-░▒╗∙▒¹╗∙)░Ęī”╗ņ║ŽŠ·╚║╬ó╔·╬’ĖĮų°╠žąįĄ─ė░ĒæĮY╣¹╚ńłD 1╦∙╩Š.╬ó╔·╬’Įø2.62~131 mgĪżL-1ļp(3-░▒╗∙▒¹╗∙)░Ę╠Ä└Ē12 hĖĮų°┴┐Ž┬ĮĄ┴╦5.61%~67.93%, ╠Ä└Ē24 hĖĮų°┴┐Ž┬ĮĄ┴╦4.60%~74.61%.┐╔ęŖį┌▀xČ©Ą─ā╔éĆĢrķgČ╬ī”╬ó╔·╬’ĖĮų°Ą─ęųųŲū„ė├Š∙ļSų°ļp(3-░▒╗∙▒¹╗∙)░ĘØŌČ╚╔²Ė▀Č°į÷ÅŖ.
ĪĪĪĪłD 1
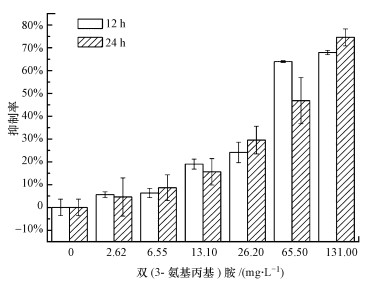
ĪĪĪĪłD 1▓╗═¼ØŌČ╚ļp(3-░▒╗∙▒¹╗∙)░Ęī”╗ņ║ŽŠ·┬õ╔·╬’─żą╬│╔ė░Ēæ ĪĪĪĪ3.2 ▓╗═¼╗∙Ąūīė╔Žļp(3-░▒╗∙▒¹╗∙)░Ęī”╬ó╔·╬’ĖĮų°ė░Ēæ
ĪĪĪĪ╬ó╔·╬’ĖĮų°ĄĮ▌d¾w▒Ē├µ╩Ū╔·╬’─żą╬│╔Ą─Ą┌ę╗▓Į, ┐╔ęį═©▀^╠ž«Éąį╗“š▀ĘŪ╠ž«ÉąįĄ─╝ÜŠ·-▒Ē├µŽÓ╗źū„ė├░l╔·(Hochbaum et al., 2011).ę╗ą®▒Ē├µ╠žąįĢ■ė░Ēæ╬ó╔·╬’Ą─ĖĮų°, Ūęę╗ą®ęųųŲä®ī”╬ó╔·╬’ĖĮų°ęųųŲū„ė├ę▓Ģ■╩▄ĄĮ▒Ē├µ╠žąįĄ─ė░Ēæ.ę“┤╦, ▒ŠčąŠ┐ī”ė┌▓╗═¼╗∙Ąūīė╔Žļp(3-░▒╗∙▒¹╗∙)░Ęī”╬ó╔·╬’ĖĮų°ė░Ēæ╠žąį▀Mąą┐╝▓ņ.ļp(3-░▒╗∙▒¹╗∙)░Ę┤µį┌Ą─ŪķørŽ┬, ĖĮų°ĄĮ╗∙Ąū▓─┴ŽĄ─╔·╬’┴┐ĮøĮYŠ¦ūŽ╚Š╔½ėøõø, ╚ńłD 2╦∙╩Š.ĮY╣¹’@╩Š¤ošō╩Ūį┌╩Ķ╦«ąįĄ─Š█▒Įęꎮ▒Ē├µ▀Ć╩ŪėH╦«ąįĄ─Š█╝ū╗∙▒¹Ž®╦ß╝ū§ź║═▓Ż┴¦▒Ē├µ, ļp(3-░▒╗∙▒¹╗∙)░Ęī”╬ó╔·╬’Ą─ĖĮų°Š∙ėą║▄║├ĄžęųųŲū„ė├.šf├„▀@ĘN╬’┘|┐╔ęįÅVĘ║æ¬ė├ė┌▓╗═¼▓─┘|▓─┴Ž▒Ē├µ╔·╬’─żĄ─┐žųŲ.
ĪĪĪĪłD 2

ĪĪĪĪłD 2▓╗═¼╗∙Ąūīė▒Ē├µļp(3-░▒╗∙▒¹╗∙)░Ęī”╬ó╔·╬’ĖĮų°ė░Ēæ
ĪĪĪĪ3.3 ļp(3-░▒╗∙▒¹╗∙)░Ęī”─ż╬█╚ŠŠÅĮŌą¦æ¬čąŠ┐
ĪĪĪĪ┴Ē═Ō, ×ķ┴╦▀Mę╗▓Į╠Įėæļp(3-░▒╗∙▒¹╗∙)░Ę┐žųŲ─ż▒Ē├µ╔·╬’╬█╚ŠĄ─┐╔─▄ąį, īóļp(3-░▒╗∙▒¹╗∙)░Ę═Č╝ėĄĮ╚▄ę║═©▀^║═æęĖĪ╬ó╔·╬’ū„ė├ų«║¾, ┐╝▓ņ╬ó╔·╬’į┌─ż▒Ē├µĄ─ĖĮų°ą╬æB, ĘųäeęįņoæBĖĮų°║═äėæB▀^×Vą╬╩Į┐╝▓ņ.ņoæBą╬╩Į┼ÓBŚl╝■Ž┬, ╬ó╔·╬’Įø▀^▓╗═¼ØŌČ╚Ą─ļp(3-░▒╗∙▒¹╗∙)░Ęū„ė├║¾į┌Š█§Ż░Ę─ż▒Ē├µĄ─ĖĮų°┴┐╚ńłD 3╦∙╩Š.ĮY╣¹’@╩Š, ╬ó╔·╬’Įø▀^ļp(3-░▒╗∙▒¹╗∙)░Ę╠Ä└Ēį┌─ż▒Ē├µĄ─ĖĮų°┴┐├„’@ĮĄĄ═, Č°Ūęļp(3-░▒╗∙▒¹╗∙)░ĘØŌČ╚įĮĖ▀, ĖĮų°┴┐įĮ╔┘.└²╚ń, Įø▀^26.2Īó65.5╝░131.0 mgĪżL-1ļp(3-░▒╗∙▒¹╗∙)░Ę╠Ä└Ē║¾, ╬ó╔·╬’į┌─ż▒Ē├µĖĮų°┴┐ĘųäeĮĄĄ═┴╦18.38%Īó31.03%║═69.15%.ļp(3-░▒╗∙▒¹╗∙)░Ę’@ų°ęųųŲ┴╦╬ó╔·╬’į┌─ż▒Ē├µĄ─ĖĮų°, Č°Ūę▀@ĘNęųųŲą¦╣¹║═ļp(3-░▒╗∙▒¹╗∙)░ĘØŌČ╚│╔š²ŽÓĻP.┤╦ĮY╣¹ę▓į┌SEMųąĄ├ĄĮūCīŹ, ī”ššĮM─ż▒Ē├µą╬│╔Ą─╬ó╔·╬’╚║┬õ▒╚▌^ų┬├▄, ╔·╬’─ż▀B│╔Ų¼, Č°īŹ“×ĮM─ż▒Ē├µĖĮų°Ą─╬ó╔·╬’ŽÓ«öŽĪ╩Ķ, ėą┤¾┴┐Ą─┐š░ūģ^ė“, īŹ“×ĮM─ż▒Ē├µĖĮų°Ą─╔·╬’┴┐ę¬├„’@╔┘ė┌ī”ššĮM─ż▒Ē├µĖĮų°Ą─╬ó╔·╬’┴┐.
ĪĪĪĪłD 3
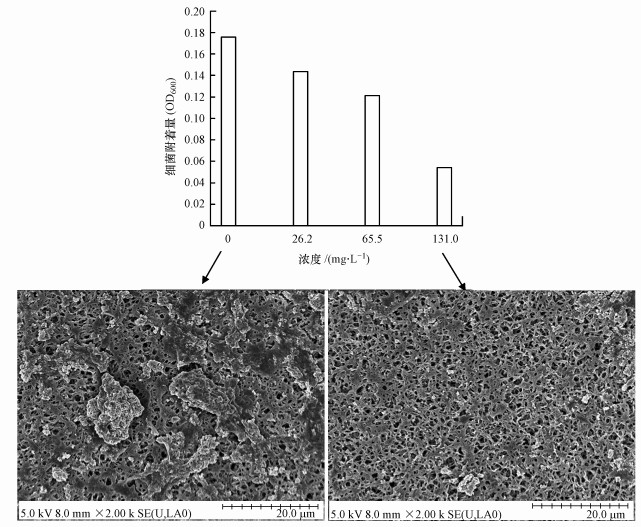
ĪĪĪĪłD 3ļp(3-░▒╗∙▒¹╗∙)░Ęī”╗ņ║ŽŠ·┬õ╬ó╔·╬’į┌Š█§Ż░Ę─ż▒Ē├µĄ─ĖĮų°ė░Ēæ
ĪĪĪĪī”ė┌╔·╬’─żäėæB▀^×V┼ÓB▀^│╠, ─ż▀^×V▀^│╠ųą─żē║ļS▀^×Vę║┘|┴┐╝░Ģrķgūā╗»╚ńłD 4╦∙╩Š.ė╔łD┐╔ęį┐┤│÷, «ö▀^×Vųž┴┐×ķ359.71 g, ▀^×VĢrķg170 minĢr, ī”ššĮM─ż┐ū(╝┤▀^×Vę║ųąø]ėąļp(3-░▒╗∙▒¹╗∙)░Ę)░l╔·┴╦ę╗Č©│╠Č╚Ą─Č┬╚¹, ┤╦Ģr─żē║×ķ10 kPa, Ą½īŹ“×ĮM(╝┤▀^×Vę║ųą╝ėėąļp(3-░▒╗∙▒¹╗∙)░Ę)ųąĄ──żē║╚į╚╗×ķ0, ▒Ē├„─ż┐ūÅĮ▓óø]ėąŽ±ī”ššĮM─żČ┬╚¹ć└ųž;«ö▀^×Vę║┘|┴┐×ķ628.55 g, ▀^×VĢrķg×ķ300 minĢr, īŹ“×ĮMųąĄ──żē║┴”ķ_╩╝╔Ž╔², ─żē║×ķ12.5 kPa, ╝s×ķ▀^×VĢrķg175 minĢrĄ─ī”ššĮM─żē║, Č°┤╦Ģrī”ššĮMųąĄ──żē║ęčĮø×ķ50 kPa.▀@▒Ē├„ļp(3-░▒╗∙▒¹╗∙)░Ę┐╔£pŠÅ─ż▀^×V▀^│╠ųą─ż┐ūČ┬╚¹╦┘Č╚, ĮĄĄ═─żē║, ŠÅĮŌė╔╬ó╔·╬’ę²ŲĄ──ż╬█╚Šå¢Ņ}.
ĪĪĪĪłD 4
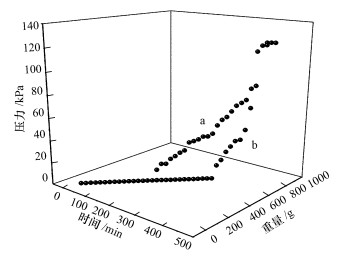
ĪĪĪĪłD 4─ż▀^×V▀^│╠─żē║ļS▀^×V┘|┴┐║═▀^×VĢrķgūā╗»łD (a.ī”ššĮM; b.īŹ“×ĮM)
ĪĪĪĪ3.4 ļp(3-░▒╗∙▒¹╗∙)░Ęī”╗ņ║ŽŠ·╚║╔·╬’─żĮŌ¾wė░Ēæ
ĪĪĪĪę╗ą®é„ĮyĄ─╬’└Ē╗»īWĘĮĘ©, ╚ńø_╦óĪóŽ¹ČŠä®╝░ūŽ═ŌŠĆŽ¹ČŠĄ╚│Ż▒╗ė├ū„╔·╬’─ż╚ź│², Ą½ė╔ė┌╔·╬’─żųą╝ÜŠ·╩▄EPSĄ─▒Żūoū„ė├╩╣Ųõī”▀@ą®│ŻęÄĘĮĘ©Š▀ėąę╗Č©┐╣ąį, ╝┤╩╣Ė▀ØŌČ╚Ą─ÜóŠ·ä®ę▓║▄ļyīó╔·╬’─ż╚ź│².└²╚ń, 500 mgĪżL-1Ą─│ŻęÄäāļxä®ę▓ļyęįäāļx│╔╩ņĄ─╔·╬’─ż, ūŽ═ŌŠĆī”╔·╬’─żųą╬ó╔·╬’Üó£ń─▄┴”├„’@╚§ė┌ī”æęĖĪ╬ó╔·╬’Ą─Üó£ńū„ė├(═§║ŻĘÕĄ╚, 2010).▒ŠčąŠ┐▀Mę╗▓Įī”ąĪĘųūė╬’┘|ļp(3-░▒╗∙▒¹╗∙)░Ę─▄ʱę²Ų│╔╩ņ╔·╬’─żĮŌ¾w▀Mąą╠Įėæ.131.0 mgĪżL-1ļp(3-░▒╗∙▒¹╗∙)░Ęī”ė┌ŅA┼ÓB12 hĄ─╔·╬’─żĮŌ¾wą¦╣¹╚ńłD 5╦∙╩Š.ė╔łDĄ├│÷, ļp(3-░▒╗∙▒¹╗∙)░Ę╠Ä└Ē╔·╬’─ż6~12 h║¾, ╔·╬’─ż┴┐ŽÓī”ė┌ī”ššĮMŽ┬ĮĄ┴╦3.87%~23.59%, ė╔┤╦┐╔ęįšf├„ļp(3-░▒╗∙▒¹╗∙)░Ęī”╔·╬’─żĮŌ¾wėąę╗Č©Ą─┤┘▀Mū„ė├.▀@ī”ė┌┐žųŲ╔·╬’─ż, ĮĄĄ═╬ó╔·╬’░▓╚½’LļUŠ▀ėąųžę¬Ą─ęŌ┴x.Š▀¾w┬ōŽĄ╬█╦«īÜ╗“ģóęŖhttp://www.jianfeilema.cnĖ³ČÓŽÓĻP╝╝ąg╬─ÖnĪŻ
ĪĪĪĪłD 5
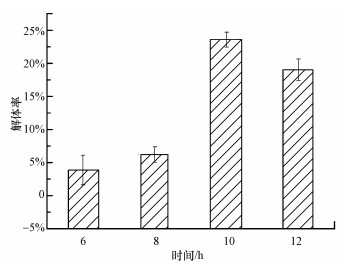
ĪĪĪĪłD 5ļp(3-░▒╗∙▒¹╗∙)░Ęī”│╔╩ņ╔·╬’─żĄ─ĮŌ¾wū„ė├
ĪĪĪĪ3.5 ļp(3-░▒╗∙▒¹╗∙)░Ęī”╬ó╔·╬’EPSĘų├┌Ą─ė░Ēæ
ĪĪĪĪ×ķ┴╦▀Mę╗▓Į╠ĮŠ┐ļp(3-░▒╗∙▒¹╗∙)░Ęī”╬ó╔·╬’Ą─ū„ė├ÖCųŲ, ▒ŠčąŠ┐▀Mę╗▓Į┐╝▓ņ┴╦ļp(3-░▒╗∙▒¹╗∙)░Ęī”╬ó╔·╬’╔·ķL╝░ŲõEPS(░¹═ŌĄ░░ūĪóČÓ╠Ū║═DNA)Ą─ė░Ēæ.ĮY╣¹╚ńłD 6╦∙╩Š, 131 mgĪżL-1ļp(3-░▒╗∙▒¹╗∙)░Ęī”╬ó╔·╬’12 h╔·ķL┴┐Ą─ęųųŲ┬╩×ķ0.36%Ī└0.54%, ø]ėąęųųŲ╬ó╔·╬’╔·ķL;ī”ššĮMųą░¹═ŌĄ░░ūĪóČÓ╠Ū║═eDNAĄ─║¼┴┐Ęųäe╩Ū(8.48Ī└0.46) mgĪżg-1 MLVSSĪó(3.04Ī└0.20) mgĪżg-1 MLVSS║═(18.33Ī└2.55) ”╠gĪżg-1 MLVSS, Įø▀^ļp(3-░▒╗∙▒¹╗∙)░Ę╠Ä└Ēų«║¾, ░¹═ŌČÓ╠Ū║═eDNAŠ∙ėąŽ┬ĮĄ, Ęųäe×ķ(1.85Ī└0.08) mgĪżg-1 MLVSS, (5.49Ī└0.54) ”╠gĪżg-1 MLVSS, Ž┬ĮĄ┴╦39.37%Ī└2.68%║═70.05%Ī└2.93%, Č°░¹═ŌĄ░░ū║¼┴┐Äū║§ø]ėąūā╗», Š∙▒Ż│ųį┌8.02~8.98 mgĪżg-1 MLVSS.▒ŠšnŅ}ĮM▀Mę╗▓Į═©▀^╝ż╣Ō╣▓Š█Į╣ė^▓ņ╝░Č©┴┐ėŗ╦Ń’@╩Šļp(3-░▒╗∙▒¹╗∙)░Ęī”╔·╬’─ż╠Ä└Ē║¾╩╣ČÓ╠Ū£p╔┘┴╦53%(Wu et al., 2016).ė╔┤╦┐╔ęį═ŲöÓļp(3-░▒╗∙▒¹╗∙)░Ęī”╬ó╔·╬’ĖĮų°╝░╔·╬’─żą╬│╔Ą─ęųųŲū„ė├▓ó▓╗╩Ū═©▀^ÜóŠ·«a╔·, ī”╬ó╔·╬’EPSųą░¹═ŌČÓ╠Ū║═eDNAĘų├┌ėąę╗Č©Ą─ęųųŲū„ė├, Ą½ī”░¹═ŌĄ░░ūø]ėąė░Ēæ.
ĪĪĪĪłD 6
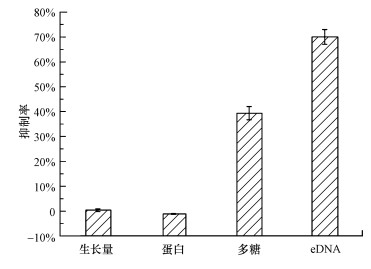
ĪĪĪĪłD 6ļp(3-░▒╗∙▒¹╗∙)░Ęī”╬ó╔·╬’╔·ķL╝░EPS«a┴┐Ą─ė░Ēæ
ĪĪĪĪō■ł¾Ą└, ļp(3-░▒╗∙▒¹╗∙)░ĘĮYśŗųą║¼ėąĄ─░▒╗∙┐╔ęįų▒Įė║═ę╗ą®Ä¦žōļŖ║╔Ą─╗∙łF╗“š▀╩Ū░¹═ŌČÓ╠Ūųąę╗ą®Ä¦ėąśOąį╗∙łFĄ─╠ŪŅÉŽÓ╗źū„ė├(Kropec et al., 2005; Sutherland, 2001), ▀@┐╔─▄╩ŪŲõę²ŲČ■š▀ØŌČ╚ĮĄĄ═Ą─ųžę¬įŁę“.▒Ŗ╦∙ų▄ų¬, EPS(Ą░░ūĪóČÓ╠Ū║═eDNA)ī”ė┌╬ó╔·╬’ĖĮų°╝░Ųõ╔·╬’─żą╬│╔Ųų°ų┴ĻPųžę¬Ą─ū„ė├, ╦³┤┘╩╣╬ó╔·╬’į┌╔·╬’║═ĘŪ╔·╬’▒Ē├µĄ─│§╩╝ĖĮų°.īóEPS═Ļ╚½╚ź│²ęį║¾, ╬ó╔·╬’Ą─ĖĮų°ąį─▄’@ų°ĮĄĄ═.░¹═ŌČÓ╠Ū╩Ū╔·╬’─żą╬│╔Ą─šTī¦ę“ūė, ┐╔╩╣▒Ē├µŅA╠Ä└Ē╗»ĖĮų°Ė³╚▌ęū░l╔·, ╔§ų┴┐╔ęįū„×ķ╝ÜŠ·╠╝į┤(Beauregard et al., 2013);eDNAī”ė┌╬ó╔·╬’į┌ėH╦«ąį║═╩Ķ╦«ąį▒Ē├µĄ─ĖĮų°Č╝Ųų°ųžę¬Ą─ū„ė├(Das et al., 2010), ╦³═©▀^ČÓĘNĘĮ╩Įš{┐ž╔·╬’─żĄ─░lš╣▀^│╠, ╚ń×ķŲõ╦¹╝ÜŠ·╠ß╣®╗∙┘|Īó╦³╩Ū╔·╬’─żĮYśŗĄ─ųžę¬│╔ĘųĪó┐╔┤┘▀M╗∙ę“╬’┘|Ą─Į╗ōQ(Wu et al., 2009)╝░š{┐žĮY║Žąįöz╚ĪŽĄĮyĄ╚(Petersen et al., 2005).┴Ē═Ō, EPSųąĄ─Ą░░ūĮM┤¾ČÓ×ķĦš²ļŖ║╔Ą─╩Ķ╦«ąį╬’┘|ĪóČÓ╠Ū×ķĦžōļŖ║╔Ą─ėH╦«ąį╬’┘|╝░eDNA×ķĦžōļŖ║╔Ą─╩Ķ╦«ąį╬’┘|(Sheng et al., 2010), ╚²š▀ų«ķgĄ─ŽÓ╗źū„ė├ŠS│ų┴╦╝ÜŠ·Ą─▒Ē├µ╠žąį, «ö▀@╚²š▀ĮM│╔░l╔·ūā╗»ĢrĢ■įņ│╔╝ÜŠ·▒Ē├µ╠žąįĄ─ūā╗», Č°╝ÜŠ·▒Ē├µ╠žąį(ėH╩Ķ╦«ąį║═▒Ē├µļŖ║╔)║═╬ó╔·╬’Ą─ĖĮų°├▄ŪąŽÓĻP(Harimawan et al., 2011).╦∙ęį▒ŠčąŠ┐ųąļp(3-░▒╗∙▒¹╗∙)░Ę═©▀^ęųųŲ╬ó╔·╬’ųąEPS─│ą®│╔ĘųĄ─╠žąį╗“š▀Ęų├┌, ▀MČ°ė░Ēæ╬ó╔·╬’ĖĮų°║═╔·╬’─żą╬│╔, ▓óŪęī¦ų┬ę╗ąĪ▓┐ĘųĄ─╔·╬’─ż┐╔ęįÅ─EPSĄ─░³╣³ųąßīĘ┼│÷üĒĮŌ¾w×ķæęĖĪæB╝ÜŠ·, ╝┤ę²Ų╔·╬’─żĮŌ¾w.ęčėąę╗ą®ł¾Ą└ę▓░l¼F«öEPSųąĄ░░ūĪóČÓ╠Ū║═eDNAØŌČ╚ĮĄĄ═Ģr, ╬ó╔·╬’ĖĮų°┴┐ŽÓ欎┬ĮĄ(Lv et al., 2014; Xu et al., 2011).
ĪĪĪĪ4 ĮYšō(Conclusions)
ĪĪĪĪ1) ļp(3-░▒╗∙▒¹╗∙)░Ę─▄ėąą¦ęųųŲ╗ņ║ŽŠ·╚║╬ó╔·╬’ĖĮų°║═╔·╬’─żą╬│╔, ļSų°ØŌČ╚Ą─╔²Ė▀ęųųŲū„ė├į÷ÅŖ, ŪęŲõęųųŲą¦╣¹▌^×ķĘĆČ©, ęųųŲ┬╩▓╗Ģ■ļSų°ĢrķgčėķLČ°ĮĄĄ═.
ĪĪĪĪ2) ļp(3-░▒╗∙▒¹╗∙)░Ęī”│╔╩ņ╔·╬’─żĮŌ¾wėąę╗Č©Ą─┤┘▀Mū„ė├.▀@ī”ė┌┐žųŲ╔·╬’─ż, ĮĄĄ═╬ó╔·╬’░▓╚½’LļUŠ▀ėąųžę¬Ą─ęŌ┴x.
ĪĪĪĪ3) ļp(3-░▒╗∙▒¹╗∙)░Ęį┌╔·╬’─żą╬│╔ėąą¦ęųųŲØŌČ╚(26.2~131.0 mgĪżL-1)ĘČć·ā╚▓óø]ėąī”ė┌╬ó╔·╬’╔·ķL«a╔·’@ų°ū„ė├, ę“┤╦▓╗╩Ū═©▀^ÜóŠ·ĘĮ╩ĮęųųŲ╔·╬’─żą╬│╔, Č°╩Ū═©▀^ęųųŲ╬ó╔·╬’ųąEPS║¼┴┐, ┐╔ęį▒▄├Ōé„ĮyÜóŠ·ä®«a╔·Ą─┐╣╦Äąįå¢Ņ}, ę“┤╦×ķ╔·╬’─żęųųŲ╠ß╣®┴╦ą┬ĘĮĘ©.
ĪĪĪĪ4) ļp(3-░▒╗∙▒¹╗∙)░Ę┐╔£pŠÅ─ż▀^×V▀^│╠ųą─ż┐ūČ┬╚¹╦┘Č╚, ĮĄĄ═─żē║, ŠÅĮŌė╔╬ó╔·╬’ę²ŲĄ──ż╬█╚Šå¢Ņ}, ▀@ī”ŁhŠ│ųą─ż╬█╚Š┐žųŲŠ▀ėąØōį┌Ą─ųĖī¦ęŌ┴x.(üĒį┤Ż║ŁhŠ│┐ŲīWīWł¾ ū„š▀Ż║╦ŠąŃśs)

